Přehled metabolismu aminokyselin
Aminokyseliny jsou v organismu převážně vázány v proteinech. Volné aminokyseliny vytvářejí aminokyselinovou hotovost („pool“), která je využívána k syntéze proteinů a dalších biologicky aktivních molekul (např. hormonů, neurotransmiterů, hemu, kreatinu, purinů; viz. Kapitola 11). Pokud příjem aminokyselin do poolu převáží potřeby organismu, jsou nadbytečné aminokyseliny zpracovány v intermediárním metabolismu až na vodu, oxid uhličitý a močovinu.
Katabolismus aminokyselin téměř vždy začíná odstraněním aminoskupiny. Ta se buď uvolní ve formě amoniaku, nebo se přenese na vhodný akceptor, kterým je 2-oxokyselina (např. pyruvát, oxalacetát, oxoglutarát). Druhá reakce (transaminace) se týká většího počtu aminokyselin. Transaminace je katalyzována aminotransferasami, které využívají jako koenzym pyridoxal-5‘-fosfát, derivát vitaminu B6 pyridoxinu. Reakce probíhá ping-pongovým mechanismem, kdy je nejprve z transaminované aminokyseliny přenesena aminoskupina na koenzym a vzniká oxokyselina. Ve druhé fázi reakce je aminoskupina z koenzymu přenesena na akceptor (2-oxokyselinu) a vzniká nová aminokyselina. Příkladem může být transaminace aminokyseliny aspartátu, při které vzniká oxalacetát a aminoskupina je připojena na akceptor 2-oxoglutarát, z něhož vzniká glutamát. Díky tomu, že rovnovážná konstanta transaminační reakce je blízká jedné, je reakce vratná a může sloužit i k syntéze aminokyselin.
V organismu je transaminace často spojena s oxidační deaminací, a pak se tento proces označuje jako transdeaminace. Při oxidační deaminaci je aminoskupina aminokyseliny uvolněna ve formě amoniaku. Reakci katalyzuje mitochondriální glutamátdehydrogenasa s koenzymem NAD+. Tento enzym deaminuje glutamát na 2‑oxoglutarát a amoniak, glutamátdehydrogenasa však může katalyzovat i syntézu glutamátu, ale v tomto případě použije jako koenzym NADPH. Deaminační reakce se výrazně urychlují při dlouhodobém hladovění a mohou vznikat až toxické koncentrace amoniaku. Dalším enzymem schopným katalyzovat oxidační deaminaci je oxidasa L-aminokyselin, která je přítomná jen v peroxisomech buněk jater a ledvin.
Amoniak je pro buňky toxický a musí být proto detoxikován. Nejcitlivější k jeho působení jsou buňky mozku. Lidský organismus přeměňuje toxický amoniak převážně v močovinovém cyklu (pouze játra) na netoxickou močovinu, která je dobře rozpustná ve vodě a snadno difunduje (viz. dále). Některé buňky, zejména mozkové, detoxikují amoniak okamžitě. Glutaminsyntetasa v těchto buňkách připojuje vznikající amoniak k molekule glutamátu za vzniku glutaminu a spotřeby ATP. Glutamin je netoxickým nosičem amoniaku a v játrech a ledvinách je z něj amoniak glutaminasou odštěpen. V ledvinách je amoniak vyloučen ve formě NH4+ močí, zatímco v játrech je detoxikován v močovinovém cyklu.
Uhlíková kostra aminokyselin se zapojuje do intermediárního metabolismu prostřednictvím společných intermediátů, které jsou součástí citrátového cyklu (Obr. 1). Tyto intermediáty se mohou napojovat na metabolismus ostatních živin i na syntézu neesenciálních aminokyselin. Uhlíková kostra aminokyselin může sloužit k syntéze glukosy (glukogenní aminokyseliny), nebo ketolátek a mastných kyselin (ketogenní aminokyseliny – poskytují acetoacetát, nebo acetyl-CoA).
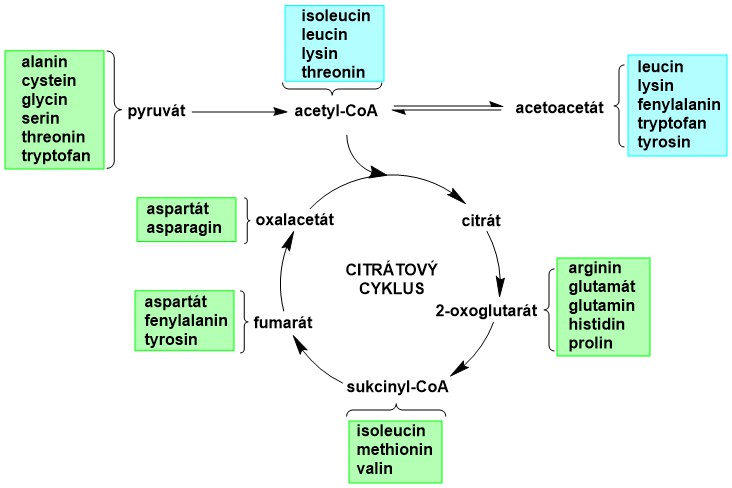
Obr. 1. Zapojení aminokyselin do intermediárního metabolismu
V dalším textu se zaměříme jen na vybrané poruchy močovinového cyklu, poruchy metabolismu či transportu aromatických, větvených a sirných aminokyselin a krátce se zmíníme o poruchách metabolismu lysinu a tryptofanu, které jsou sledovány v rámci novorozeneckého screeningu v ČR.