Trávení proteinů a vstřebávání aminokyselin v GIT
Žaludek
Trávení proteinů v GIT je zahájeno v žaludku působením žaludeční šťávy. Složení žaludeční šťávy závisí na příjmu potravy. Nalačno je v žaludku přítomno jen malé množství sekretu se slabě kyselým pH a vysokým obsahem Na+, zatímco po příjmu potravy se značně zvýší sekrece, pH se sníží do silně kyselé oblasti a obsah Na+ se sníží. Žaludeční šťáva vzniká spojením sekretu parietálních (krycích) buněk a sekretu hlavních buněk. Parietální buňky produkují silně kyselý sekret obsahující HCl (pH < 1) a vnitřní faktor, což je protein nutný pro absorpci kobalaminu (vitaminu B12). Hlavní buňky tvoří slabě zásaditý sekret obsahující pepsinogen.
Kyseliny chlorovodíková, která vzniká působením enzymu karboanhydrasy (karbonátdehydratasy)[1]. Tento enzym katalyzuje interkonverzi rozpuštěného oxidu uhličitého a hydrogenuhličitanového iontu. aktivuje v žaludeční šťávě pepsinogen na pepsin, denaturuje proteiny potravy a tím usnadňuje jejich proteolýzu, působí antibakteriálně a umožňuje solubilizaci nerozpustných Ca2+ a Fe2+ solí. Její sekrece je regulována nervově, endokrinně, parakrinně a autokrinně. Nejvýznamnější stimulační účinek má gastrin, který aktivuje protonovou pumpu H+/K+-ATPasu. Tento hormon pochází z G-buněk antra žaludku Stimulační účinek má též acetylcholin, neurotransmiter postgangliových vláken nervus vagus, který působením na M1‑receptory a neurony stimuluje výdej gastrinu prostřednictvím GRP („gastrin-releasing peptide“). Parakrinní stimulační vliv má histamin produkovaný ECL-buňkami („enterochromaffin-like“) a mastocyty žaludeční stěny. Inhibičně působí sekretin (endokrinně) z tenkého střeva, somatostatin (parakrinně) produkovaný D‑buňkami žaludeční sliznice a prostaglandiny (zejm. PGI2 a PGE2), transformující růstový faktor-α a adenosin (všechny para- a autokrinně). Významným regulačním mechanismem je zpětnovazebné tlumení sekrece gastrinu vysokou koncentrací H+ iontů v dutině žaludku (Obr. 4).
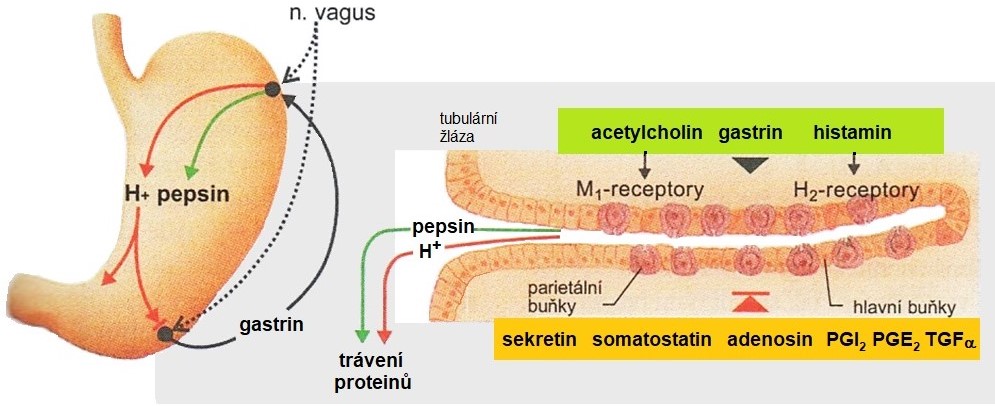
Obr. 4. Tvorba žaludeční šťávy a její regulace (upraveno Silbernagl a Lang 2012)
Druhou složkou žaludeční šťávy je pepsinogen syntetizovaný hlavními buňkami přímo úměrně rychlosti sekrece HCl. Působením HCl dojde k aktivaci části molekul pepsinogenu na pepsin, aktivace dalších molekul probíhá autokatalyticky. Pepsin je málo specifická endopeptidasa s pH optimem v silně kyselé oblasti, pro jejíž katalytickou aktivitu je rozhodující aspartátový zbytek v aktivním místě. Výsledkem působení pepsinu na proteiny potravy je směs kratších peptidů (tzv. pepton). Po promísení tráveniny (chymu) s pankreatickou šťávou, která má alkalické pH, je účinek pepsinu nevratně ukončen.
V žaludku kojenců a dalších mláďat savců se nachází proteolytický enzym chymosin (též rennin), který za účasti Ca2+ sráží kasein mléka na parakasein a tím brání rychlému průtoku mléka do tenkého střeva. Parakasein je následně štěpen pepsinem. U dospělých jedinců chymosin zcela chybí.
Tenké střevo
Vstup chymu do duodena aktivuje sekreci pankreatické šťávy, která obsahuje řadu trávicích enzymů, jejichž pH optimum se nachází v neutrální až slabě alkalické oblasti. Alkalické pH pankreatické šťávy (pH 7,4-8,3), které je způsobeno vysokou koncentrací HCO3‾, je důležité rovněž pro neutralizaci žaludeční HCl a inaktivaci pepsinu. Pankreatická šťáva je nejúčinnějším trávicím médiem, protože je díky obsahu hydrolytických enzymů schopná trávit všechny druhy živin. Tvorba pankreatické šťávy a sekrece HCO3‾ je stimulována působením hormonu sekretinu, který je produkován buňkami duodena, jejuna a částečně i buňkami žaludeční sliznice. Sekrece trávicích enzymů pankreatu a kontrakce žlučníku jsou stimulovány cholecystokininem (též pankreozymin).
Proteolytické enzymy obsažené v pankreatické šťávě jsou acinárními buňkami pankreatu produkovány ve formě zymogenů. Ty jsou aktivovány proteolytickým odstraněním části své molekuly. Mezi nejvýznamnější peptidasy pankreatické šťávy patří trypsin, chymotrypsin, elastasa a karboxypeptidasy. Sliznice tenkého střeva produkuje enzym enterokinasu (též enteropeptidasu), která aktivuje část molekul trypsinogenu na trypsin. Trypsin pak autokatalyticky aktivuje další molekuly trypsinogenu, ale též chymotrypsinogen, proelastasu a prokarboxypeptidasu (Obr. 5).
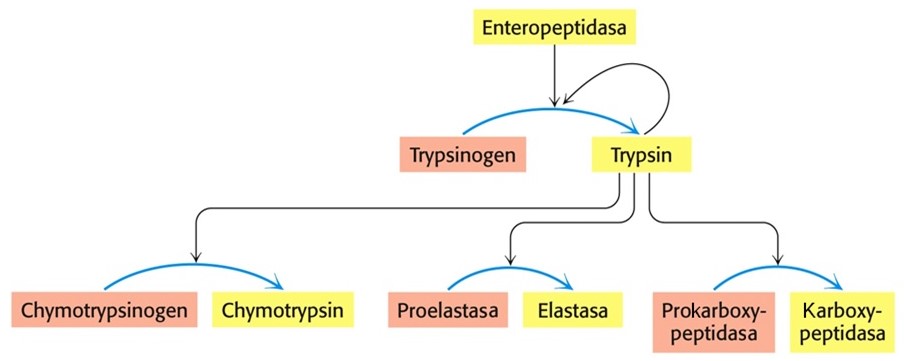
Obr. 5. Aktivace pankreatických zymogenů (upraveno z Berg et al. 2006)
Trypsin, chymotrypsin a elastasa jsou serinové endopeptidasy, zatímco karboxypeptidasa patří mezi exopeptidasy a odštěpuje poslední aminokyselinový zbytek z C-konce proteinu/peptidu. Výsledkem působení těchto enzymů je směs volných aminokyselin a oligopeptidů. Aminokyseliny jsou transportovány do enterocytů, zatímco oligopeptidy jsou na povrchu buněk kartáčového lemu doštěpeny pomocí dipeptidas a aminopeptidas. Krátké di- a tripeptidy jsou transportovány do enterocytu systémem PEPT1 a štěpeny na aminokyseliny peptidasami uvnitř těchto buněk (Obr. 6).
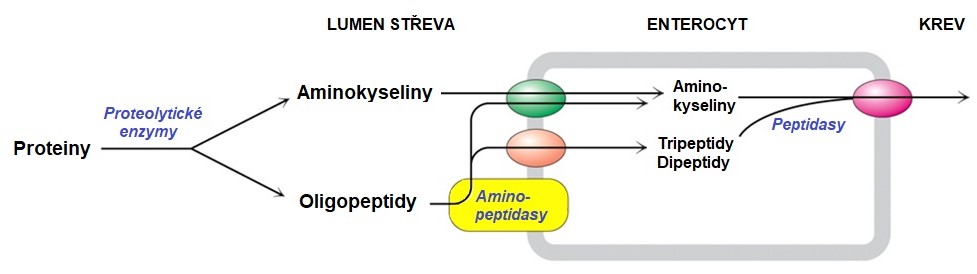
Obr. 5. Trávení proteinů v tenkém střevě a vstřebávání vznikajících produktů (upraveno z Berg et al. 2006)
Aminokyseliny vzniklé trávením proteinů v GIT jsou transportovány do enterocytů pomocí řady transportních systémů, z nichž každý přenáší skupinu příbuzných aminokyselin (neutrálních, kyselých, bazických a iminokyselin). Tento transport je aktivní (spotřeba ATP) a jsou známy Na+-dependentní a H+-dependentní transportéry (Obr. 7). Jejich umístění na povrch enterocytu se liší. Na apikální straně membrány jsou převážně zastoupeny Na+‑dependentní transportéry, které zprostředkovávají rychlý a úplný odsun aminokyselin z lumen střeva. Transportéry na bazolaterální straně membrány přenášejí aminokyseliny z enterocytu do řečiště portální žíly. Obdobným mechanismem dochází ke zpětné resorpci aminokyselin v tubulech ledvin.
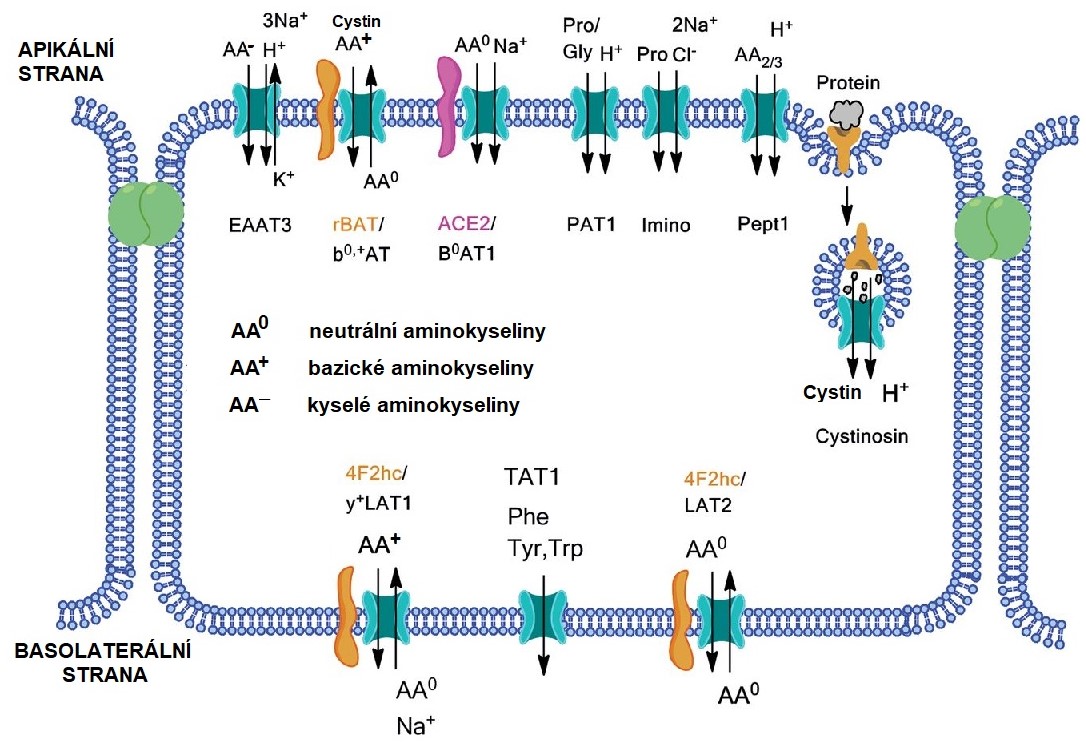
Obr. 6. Transportní systémy pro aminokyseliny ve střevě (upraveno z Bröer a Palacín 2011)
Kombinací imunochemických a elektron-optických metod bylo prokázáno, že proteiny mohou v nezměněné formě procházet ze střeva do krevního oběhu. Byly popsány dva způsoby tohoto průchodu. První se odehrává jako transcelulární přenos, kdy se proteiny vážou na membránu mikroklků, endocytosou jsou přijaty do cytoplasmy enterocytů a exocytosou vypuštěny do krve. Další možností transcelulárního přenosu je transport proteinů pomocí lymfatického systému střev. Střevní epitel obsahuje M-buňky, které transportují enzymy do lymfatické tkáně (Peyerovy plaky), tam se proteiny váží na lymfocyty, které je lymfatickým systémem dopraví až do krevního řečiště. Druhým, a významnějším, způsobem přenosu proteinů je paracelulární průnik v tzv. „zóně línání“, která vzniká při dočasném defektu po odloupnutí starých enterocytů. Skutečnosti, že proteiny mohou procházet v nezměněné formě střevní stěnou do krve, se využívá v enzymoterapii.
[1] Karboanhydrasa katalyzuje reverzibilní přeměnu oxidu uhličitého a vody na kyselinu uhličitou, která z velké části disociuje na hydrogenuhličitanový ion a vodíkový proton (CO2 + H2O ↔ H2CO3 ↔ HCO3- + H+), jehož export z buňky výměnou za jiný kation (např. Na+) vede k okyselení příslušného prostředí. Karboanhydrasa je obsažena v buňkách žaludeční sliznice, kde se podílí na tvorbě HCl. Dále se účastní acidifikace moči v ledvinných tubulech, přenosu CO2 v erytrocytech a produkce nitrooční tekutiny v oku.