Přehled metabolismu proteinů
Ve všech tkáních kontinuálně probíhá degradace a syntéza nových proteinů (obrat proteinů, Obr. 1). Proteiny potravy tvoří v této bilanci kolem 100 g, zatímco endogenní proteiny až 200 g za den. V organismu zdravého dospělého člověka je tedy denně odbouráno a nasyntetizováno kolem 300 g proteinů, což představuje zhruba 1-2 % jejich celkového množství. V rychlosti obratu jednotlivých proteinů a tkání jsou vlivem odlišné hormonální a nervové regulace a přítomnosti různých enzymů a transportérů v jednotlivých tkáních značné rozdíly. Velký obrat proteinů je typický pro tkáně, ve kterých dochází k velkým strukturním změnám, například v kosterních svalech v průběhu hladovění, nebo v děloze během těhotenství. Z celkového množství aminokyselin uvolněných při degradaci proteinů je kolem 75 % znovu využito k syntéze proteinů. Aminokyseliny nemají, na rozdíl od lipidů a sacharidů, speciální zásobní formu. Proto jsou ty, které nejsou použity k syntéze nových proteinů, velmi rychle degradovány. Jejich uhlíkové skelety jsou většinou přeměněny na amfibolické intermediáty a většina dusíku je vyloučena močí ve formě netoxické močoviny.
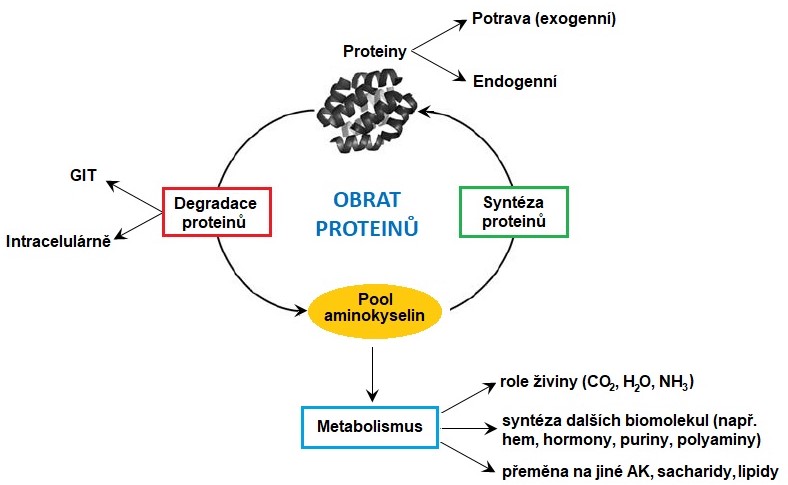
Obr. 1. Obrat proteinů a základní směry metabolismu aminokyselin (upraveno z Baskin a Taegtmeyer 2011).
K hodnocení metabolismu proteinů se v praxi využívá tzv. dusíková bilance, u které se porovnává množství přijatého a vyloučeného dusíku. U zdravých dospělých jedinců je množství dusíku přijatého potravou stejné jako množství, které se za den vyloučí (močí, stolicí, potem, ostříhanými vlasy a nehty, deskvamací rohové vrstvy epidermis) a dusíková bilance je vyrovnaná. Stav, kdy je přijato více dusíku, než je ho vyloučeno, se označuje jako pozitivní dusíková bilance a objevuje se zejména během těhotenství, růstu a rekonvalescence. Negativní dusíková bilance, tedy stav, kdy vylučování převažuje nad příjmem dusíku, vzniká v důsledku hladovění až marasmu, závažných chirurgických zákroků, rozvinutého nádorového onemocnění, nebo kwashiorkoru (závažná malnutrice vyvolaná deficitem proteinů). Dusíková bilance závisí na též příjmu esenciálních aminokyselin. Pokud chybí byť jediná esenciální aminokyselina, dusíková bilance se stává negativní a nepomůže ani nadměrná saturace dalšími aminokyselinami. Ty jsou pak jako živiny využity k získání energie a dusík je ve formě močoviny vyloučen z těla.
Proteolytické enzymy
Trávení peptidů a proteinů v trávicím traktu i jejich degradace v tkáních jsou katalyzovány peptidasami, což jsou enzymy hydrolyticky štěpící peptidovou vazbu. V GIT jsou peptidasy obsaženy v žaludeční a pankreatické šťávě a na povrchu enterocytů. Můžeme je klasifikovat podle místa hydrolýzy substrátu (endo- x exopeptidasy) a podle funkčně významných součástí enzymu (serinové, cysteinové, aspartátové a metalloproteinasy). Endopeptidasy štěpí protein kdekoliv uvnitř jeho řetězce, zatímco exopeptidasy odštěpují poslední aminokyselinový zbytek na N-konci (aminopeptidasy) nebo na C-konci (karboxypeptidasy) řetězce proteinu (Obr. 2).

Obr. 2. Rozdíl ve štěpení proteinu katalyzovaném endopeptidasou a exopeptidasou.
Na průběh štěpení proteinu má zásadní vliv několik funkčních skupin, které se nacházejí v aktivním centru peptidas. Serinové peptidasy, které v aktivním místě obsahují serin, jsou nejlépe prostudovanou třídou proteolytických enzymů. Při štěpení proteinů využívají hydroxylovou skupinu serinového zbytku k nukleofilní atace peptidové vazby. K této skupině je řazeno i několik threoninových peptidas. Serinové peptidasy jsou aktivní ve slabě alkalickém prostředí a nevyžadují žádné kofaktory. Některé enzymy jsou stabilizovány Ca2+, které se mohou účastnit aktivačních reakcí. Mezi zástupce této skupiny peptidas patří proteolytické enzymy GIT trypsin, chymotrypsin či elastasa. Tyto enzymy jsou syntetizovány ve formě inaktivních prekurzorů, zymogenů, a jsou aktivovány částečným proteolytickým sestřihem mimo syntetizující buňky. Cysteinové peptidasy mají v aktivním místě funkčně významnou thiolovou skupinu. Jedná se o převážně intracelulární enzymy a patří mezi ně kathepsiny, kalpainy, kaspasy či rostlinný papain. Aspartátové peptidasy mají v aktivním místě karboxylovou skupinu aspartátu, jsou katalyticky aktivní pouze v kyselém prostředí a patří mezi ně extracelulární i intracelulární enzymy. Mezi významné zástupce patří pepsin (extracelulární) či lyzosomální peptidasa kathepsin D. Pepsin je syntetizován ve formě neaktivního zymogenu a po aktivaci štěpí proteiny potravy v kyselém prostředí žaludku a v tenkém střevě je nevratně inaktivován pH > 7. Poslední skupinou peptidas jsou metalloproteasy, které obsahují v aktivním centru iont kovu (Zn2+, Mn2+). Tyto enzymy jsou aktivní v neutrálním prostředí a jejich aktivitu lze inhibovat chelatačními činidly. Mezi zástupce patří karboxypeptidasy pankreatické šťávy a matrix-metalloproteinasy (např. kolagenasy, gelatinasy, stromelysiny) degradující mezibuněčnou hmotu.
Peptidasy mají různou substrátovou specificitu (Obr. 3). Některé endopeptidasy, např. rostlinný papain, jsou málo specifické a štěpí řetězce na libovolném místě. Pepsin má širokou substrátovou specifitu a preferuje aromatické nebo objemné aminokyselinové zbytky, jako jsou fenylalanin (Phe), tyrosin (Tyr), tryptofan (Trp) a leucin (Leu). Trypsin je sice serinová peptidasa, ale specifitu jeho štěpení způsobuje aspartátový zbytek na dně aktivního centra, který vytváří solný můstek s bazickými aminokyselinami lysinem (Lys) a argininem (Arg), serinový a histidinový zbytek se pak podílejí na štěpení. Trypsin tedy zanechává na C-konci jednoho z produktů bazickou aminokyselinu (Lys nebo Arg). Aktivní centrum chymotrypsinu je tvořené velkou hydrofobní kapsou, takže tento enzym štěpí substrát v místě, kde se nacházejí hydrofobní aminokyseliny nebo aminokyseliny s velkými arylovými řetězci (např. Trp, Phe, Tyr, Leu, případně methionin, Met). Aktivní centrum elastasy vytváří mělkou kapsou, do které zapadnou pouze nejmenší aminokyseliny glycin (Gly) a alanin (Ala). Exopeptidasa karboxypeptidasa A odštěpuje z C-konce substrátu všechny aminokyselinové zbytky s výjimkou Arg a Lys, zatímco karboxypeptidasa B umí odstranit pouze Arg a Lys.
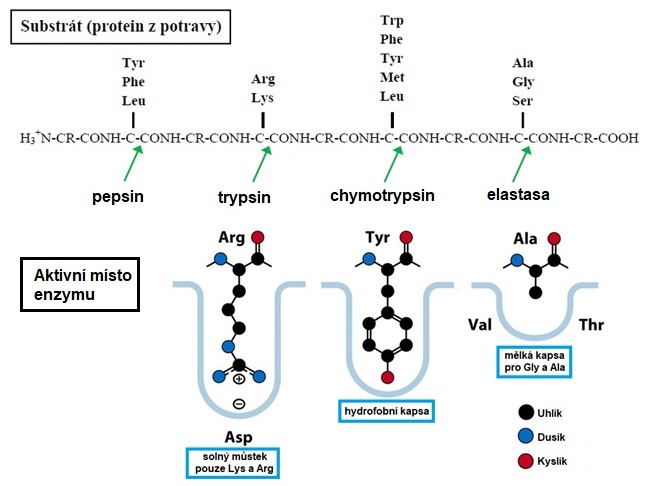
Obr. 3. Substrátová specifita a aktivní místo peptidas trávicího traktu (upraveno z Horton et al. 2006).
Prakticky všechny trávicí proteolytické enzymy vznikají aktivací zymogenů, neaktivních prekurzorů. Zymogeny jsou syntetizovány ve žlázových acinech jako velké molekuly, v Golgiho aparátu jsou pokryty membránou z proteinů a lipidů a vytvářejí zymogenní granule. Tyto granule se hromadí v apexu acinární buňky a na podnět jsou secernovány do vývodných cest. Teprve v místě potřeby jsou zymogeny aktivovány. Tímto způsobem brání organismus samonatrávení vývodných cest. Podstatou aktivace je odštěpení peptidu/ů, které maskují aktivní místo enzymu.