Metabolismus jednotlivých typů lipoproteinů
Příkladem tvorby lipoproteinů jsou chylomikra a částice VLDL, které se formují v buňkách hned v okamžiku syntézy triacylglycerolů. Díky hydrofobním vlastnostem se TAG spontánně shlukují kolem molekul esterifikovaného cholesterolu a vytváří základ lipoproteinové částice. Na povrch vznikající částice adherují strukturní apoproteiny v závislosti na tom, v které buňce se proces odehrává. V případě buněk střevní sliznice se jedná o B-48, v případě jaterních buněk o B-100. Dále se připojují další funkční apoproteiny, volný cholesterol a různé fosfolipidy (lecitiny). Konečnou podobu částice dostávají po vyloučení z buňky ven, kdy se v krvi na částici obvykle ještě napojí některé funkční apoproteiny a volný cholesterol. Molekuly apoproteinů, fosfolipidů a volného cholesterolu se natáčí tak, aby polární část jejich molekul byla směrována do vnějšího vodného prostředí. Díky hydrataci polárních skupin je pak zajištěna rozpustnost ve vodném prostředí krve. Dovnitř vznikající částice směřují hydrofobní části molekul, což vede k vytváření prostředí vhodného pro nepolární molekuly. Uvnitř částice se tedy nachází estery cholesterolu a triacylglyceroly.
Metabolismus chylomiker
V kapilárách tkání (především svalů) jsou chylomikra (jejich vznik ve střevních enterocytech a cesta do krevního řečiště byly popsány v kapitole 7) hydrolyzována lipoproteinovou lipasou na volné mastné kyseliny a glycerol. Mastné kyseliny přestupují do buněk, glycerol přechází do krve. Ve svalu je pak významné hlavně štěpení mastných kyselin β-oxidací v mitochondriích, protože slouží jako zdroj energie. Uvolněný glycerol putuje krví, je zachycován v játrech a ledvinách a využíván buď ke glukoneogenezi, nebo k syntéze triacylglycerolů. Doba životnosti chylomiker je poměrně krátká, asi 2 až 3 hodiny.
Předáním většiny tricylglycerolů buňkám se částice chylomiker zmenší a vznikají silně aterogenní částice označované jako remnanty chylomiker (zbytky chylomiker). Ty jsou metabolizovány v játrech (Obr. 5).
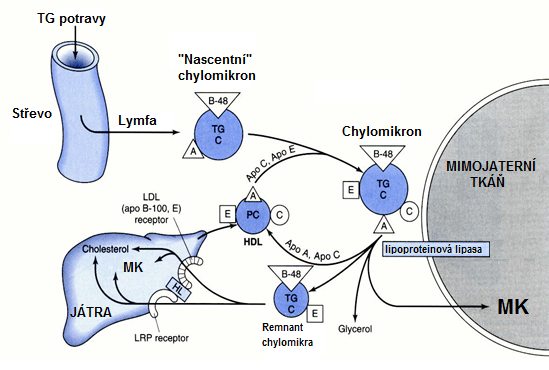
Obr. 5. Schéma vzniku a metabolismu chylomiker (upraveno z Murray et al. 2003)
Remnanty chylomiker
Chylomikra kolují krví tak dlouho, dokud se z nich neodstraní tolik triacylglycerolů, aby vznikla částice, která svou velikostí umožní transport do jaterních buněk. V remnantech se relativně vyšším úbytkem triacylglycerolů zvyšuje podíl cholesterolu, což z nich činí částice silně aterogenní. Naštěstí jsou velice rychle vychytávány játry a metabolizovány. Na povrchu hepatocytů se nachází speciální protein označovaný jako LRP (LDL receptor-related protein), který je specifickým receptorem pro apoprotein E, jenž se nachází na povrchu zbytků chylomiker (viz obr. 5). Afinita tohoto receptoru k apoproteinu E je tak vysoká, že životnost zbytkových částic chylomiker se počítá v jednotkách minut. Tím se snižuje jejich aterogenní účinek.
Vznik a metabolismus částic VLDL
Tyto lipoproteinové částice jsou syntetizovány v játrech jako transportní forma triacylglycerolů při jejich přesunu z jater do periferních tkání. Zdrojem mastných kyselin pro tvorbu triacylglycerolů jsou jednak mastné kyseliny přijaté z potravy, jednak triacylglyceroly vznikající v játrech ze sacharidů potravy. Jejich produkce závisí na příjmu sacharidů, zvyšuje se při diabetu (vysoká nabídka glukosy z krve) a také při vyšším příjmu alkoholu. U obézních osob je hranice pro iniciaci lipogeneze v játrech snížena, játra těchto jedinců snadněji vytvářejí z přijímaných sacharidů tuky.
Klíčovými proteiny pro vznik VLDL částic jsou apoproteiny A-II a B-100 (obdoba apoproteinu B-48 chylomiker v enterocytech). Významnou součástí je cholesterol, jehož zdrojem jsou remnanty chylomiker, další lipoproteinové frakce a cholesterol vzniklý endogenní syntézou v hepatocytech.
Mechanismus vytváření VLDL částic je obdobný mechanismu, který probíhá v enterocytech při vytváření chylomiker. Rozdíl je ve velikosti vzniklé částice, v obsahu cholesterolu a především v tom, že VLDL jsou transportovány z hepatocytů přímo do krve. Po uvolnění do krve se částice VLDL ještě v krvi obohacují o apoprotein E a apoproteiny C-II a C-III, které jsou přebírány od částic HDL.
Částice VLDL mohou v malé míře vznikat i v enterocytech, kde se shromažďují tuky přítomné v žluči a ty se pak transportují lymfou do krevního řečiště. Syntéza VLDL je téměř shodná se vznikem chylomikronů.
Částice VLDL jsou vylučovány z hepatocytů do krve a putují podobně jako chylomikra do cílových tkání, zejména do adipocytů. V kapilárách těchto tkání jsou jejich TAG štěpeny lipoproteinovou lipasou, lokalizovanou ve stěnách endotelu obdobně, jako je tomu u chylomiker. Lipoproteinová lipasa je přítomna jednak v tkáních, které MK používají jako palivo (např. srdce a kosterní sval), jednak v místě tvorby tukových zásob (adipocyty). Proto jsou svaly a tuková tkáň považovány za hlavní tkáně, které kontrolují koncentraci plazmatických triacylglycerolů.
Mastné kyseliny, které vstupují do tukových buněk, se především znovu esterifikují na triacylglycerol. Triacylglaceroly uložené v adipocytech jsou v případě potřeby štěpeny za katalýzy hormon-sensitivní lipasou a uvolněné mastné kyseliny jsou pak krví (ve vazbě na albuminy) transportovány např. do svalů k energetickému využití. Cíle obou lipas z hlediska krytí metabolických potřeb jsou tedy opačné a jejich hormonální regulace je proto koordinována (Obr. 6).
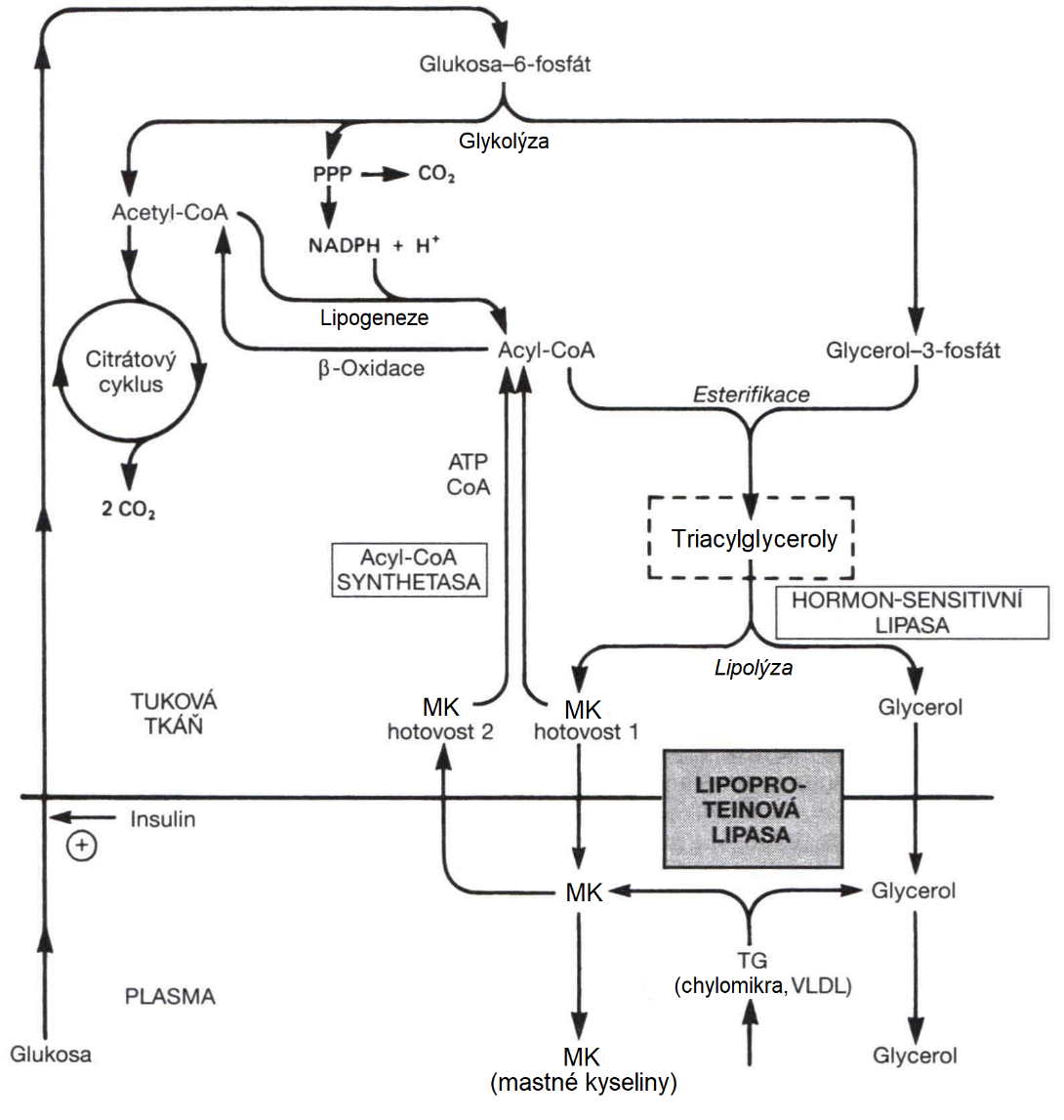
Obr. 6. Lipoproteinová lipasa a hormonsensitivní lipasa v metabolismu triacylglycerolů (Murray et al. 2001)
Snížením obsahu triacylglycerolů v částicích VLDL se částice postupně zmenší a reorganizují. Vznikají remnanty VLDL, které se běžně nazývají IDL (Intermediate Density Lipoproteins). Částice IDL jsou vychytávány játry a jsou buď zcela degradovány, nebo působením jaterní (hepatické) lipasy (HL) přeměněny na částice LDL.
IDL
Lipoproteiny o střední hustotě vznikají poté, co je z VLDL uvolněno a hydrolyzováno až 90 % triacylglycerolů, a v samotné částici zůstanou zejména molekuly cholesterolu a jeho esterů. Tyto částice jsou vysoce aterogenní, avšak jsou u zdravého člověka rychle přeměňovány na částice LDL. Jejich rizikové působení se projeví v případě, že vázne jejich metabolismus. IDL částice mají apoproteiny B-100 a E, které mají afinitu k hepatocytům. Pokud se IDL v játrech navážou na receptor LRP (pro apoprotein E), dochází k jejich degradaci. Pokud se vážou na receptor pro apoprotein B-100, jsou přeměňovány na částice LDL (Obr. 7).
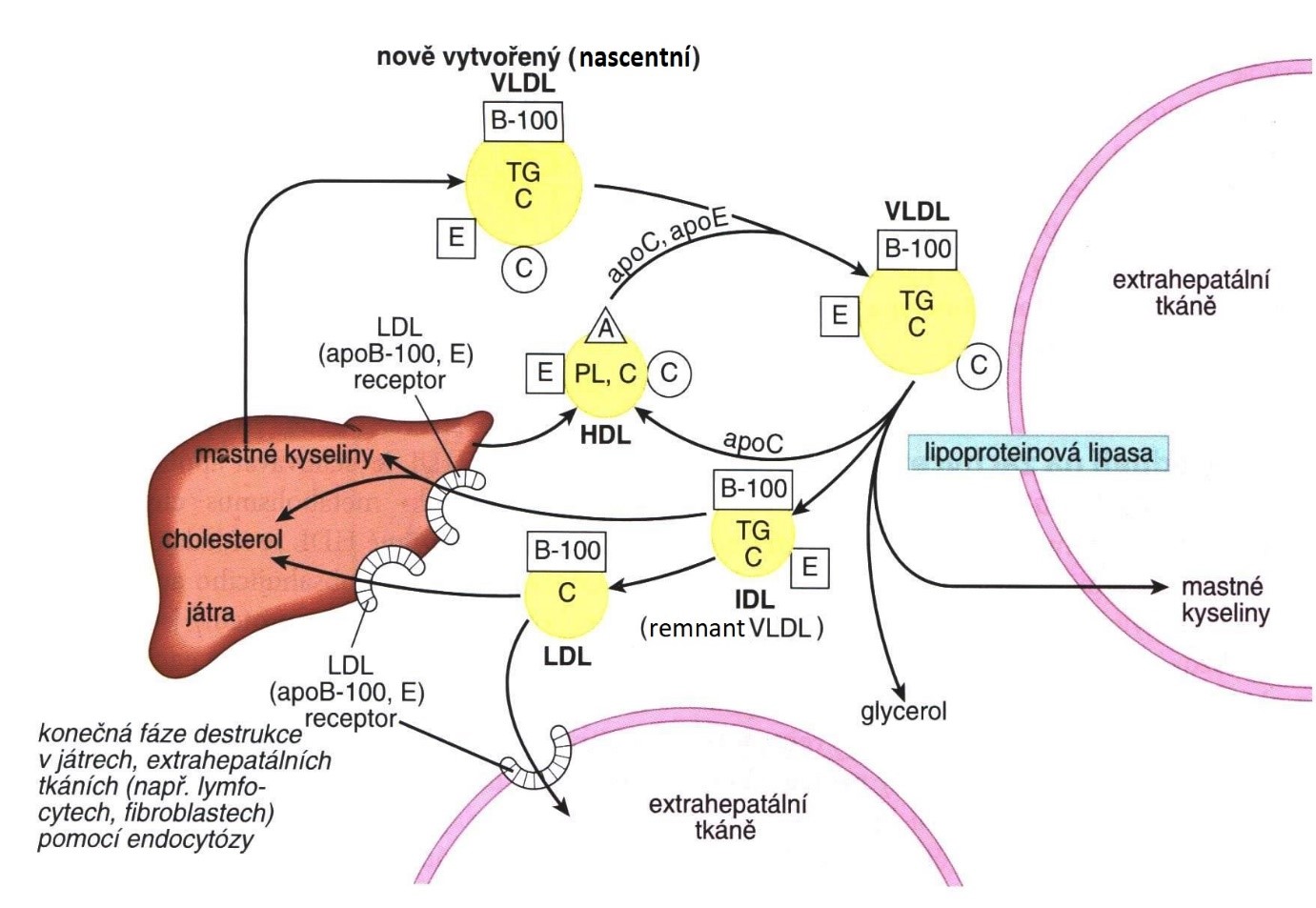
Obr. 7. Metabolismus částic VLDL, IDL a LDL (upraveno z Murray et al. 2012)
LDL
Lipoproteiny s nízkou hustotou vznikají z VLDL výše popsaným způsobem, určitá část je snad produkována přímo játry. Na rozdíl od ostatních lipoproteinových částic má LDL částice na svém povrchu lokalizovaný pouze jediný apoprotein - B-100. Funkcí LDL je dodávat cholesterol periferním tkáním. Buňky těchto tkání jsou však vybaveny i pro vlastní tvorbu cholesterolu. Výsledný efekt je pak dán souhrou příjmu cholesterolu z krve a vlastní produkcí. Poznatky o mechanismu příjmu cholesterolu z krve prostřednictvím receptoru pro apoprotein B-100 a objevení principu regulace syntézy cholesterolu v buňkách přispěly významně k porozumění úlohy cholesterolu v rozvoji aterosklerózy. J. Goldstein a M. Brown studovali v sedmdesátých letech úlohu tohoto receptoru, který se ukázal jako klíčový pro transport cholesterolu z částic LDL v krvi do buněk a hospodaření s cholesterolem (Nobelova cena v roce 1983). Při nedostatku cholesterolu začne buňka ve zvýšené míře syntetizovat LDL-receptory, které následně migrují do buněčné membrány. Po vazbě LDL částice na receptor je celý komplex procesem endocytózy vstřebán dovnitř buňky, kde dochází k rozpadu částice a k uvolnění cholesterolu. LDL-receptory se zároveň s navázáním částic LDL shlukují, takže tyto komplexy vstupují do buňky endocytózou společně ve větším kvantu. Poté se receptor uvolní z komplexu s lipoproteinem, migruje zpět směrem do membrány a celý proces se opakuje. Takto se degraduje přibližně 1/3 v periferních tkáních, zbytek v játrech. Regulace vstupu částic LDL, osudu jejich složek, receptoru pro apoprotein B-100, úlohu cholesterolu a syntézy jeho molekul v buňce de novo popisují obrázky 8 a 9.
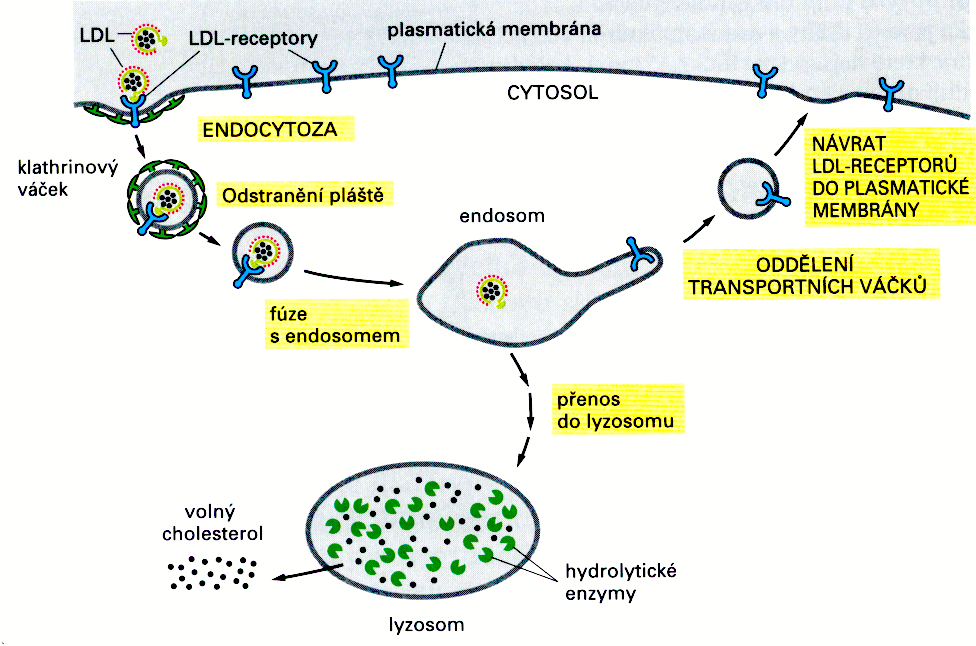
Obr. 8. Příjem částic LDL buňkou a recyklace receptorů pro LDL (upraveno z Alberts et al. 2007)
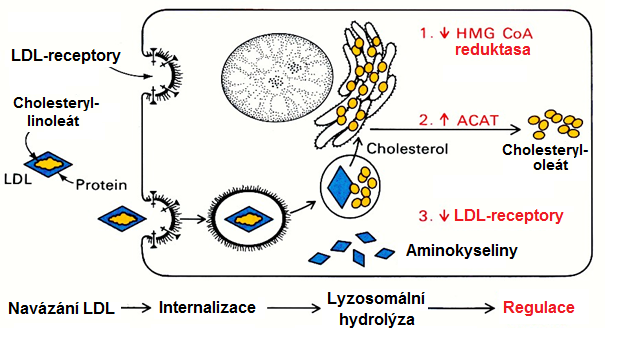
Obr. 9. Příjem částic LDL buňkou a regulace intracelulárního cholesterolu (Goldstein a Brown 2009). ACAT = Acyl-CoA cholesterolacyltransferasa (Sterol O-acyltransferasa).
LDL částice mají poměrně dlouhou dobu životnosti, během 24 hodin je katabolizováno pouze 30 až 40 % LDL, proto mohou LDL částice v krvi podléhat různým oxidačním reakcím a chemické modifikaci. Tím se snižuje afinita LDL částic k receptoru pro apoprotein B-100 na povrchu buněk a tím i jejich vstup do buněk. V této chemicky modifikované podobě vstupují (především) do makrofágů prostřednictvím tzv. scavengerových receptorů na jejich povrchu a významně přispívají k rozvoji aterosklerózy.
LDL částice jsou vysoce aterogenní a jejich míra rizikovosti vzrůstá s jejich koncentrací a se zmenšováním jejich molekul. Rizikoví jsou zejména pacienti s vrozenou hyperbetalipoproteinemií, hypertriacylglycerolemií a diabetem II. typu (viz příslušné kapitoly).
LDL částice jsou poměrně variabilní skupinou, skládající se z částic lišících se navzájem velikostí i složením. Většinou se rozlišují 4 skupiny, označované jako LDL1, LDL2, LDL3 a LDL4. Částice LDL3 a LDL4 jsou relativně menší, mají větší hustotu a jsou považovány za zvláště aterogenní.
HDL
Částice HDL vznikají v játrech a střevě dvěma mechanismy. Jednak mechanismem syntézy bílkovinného polymeru na ribosomech, přičemž dochází při vytváření terciární struktury k současnému začleňování molekul TAG, cholesterolu a fosfolipidů. Tímto způsobem vznikají nové, tzv. nascentní HDL částice diskoidního tvaru. Druhým mechanismem je shlukování uvolněných apoproteinů, fosfolipidů a cholesterolu při degradaci chylomiker příp. VLDL částic. Hlavními složkami HDL částic jsou apoproteiny A-I, A-II, C a E a dva proteiny s katalytickou aktivitou: LCAT (lecithin:cholesterolacyltransferasa) a CETP (Cholesterol Ester Transfer Protein).
Nascentní částice HDL se dostávají do kontaktu s fosfolipidy, volným cholesterolem a apoproteiny, které se uvolnily z chylomiker nebo VLDL částic (Obr. 10). Na povrchu HDL dochází k esterifikaci cholesterolu, který se díky svým hydrofobním vlastnostem přesouvá do nitra částice. Vzniká sférická částice označovaná jako HDL3. Částice HDL3 na sebe navazuje další molekuly volného cholesterolu, které opět podléhají esterifikaci a zabudování do částice. Transfer cholesterolu z buněk tkání do částice HDL se uskutečňuje pomocí transportéru ABC‑1 (ATP-binding casette transporter-1), využívajícího hydrolýzu ATP pro energetické pokrytí transmembránového transportu.
Transportem krví dochází ke kontaktu HDL3 s IDL částicemi nebo s remnanty chylomiker a k výměně triacylglycerolů za esterifikovaný cholesterol. Tento proces je podporován proteinem CETP. V důsledku toho HDL3 ztrácí část svého cholesterolu, naopak se obohacuje o triacylglyceroly. V tomto okamžiku se již mluví o částici HDL2.
Částice HDL2 vstupují do jater a jejich osud závisí na aktivitě hepatocytů. Buď jsou zachycovány pomocí receptoru, patřícího do skupiny tzv. scavengerových receptorů (SR-B1, scavengerový receptor B1) a po vstupu částice do hepatocytu dochází k její degradaci a k úplnému zániku. Její složky jsou využity pro tvorbu VLDL, případně nových nascentních HDL částic. Pokud jsou částice HDL2 atakovány jaterní lipasou, dochází k uvolnění triacylglycerolů, vznikne znovu HDL3. Tento mechanismus je upřednostněn především v období hladovění.
Základní funkce HDL spočívá v zachycování neesterifikovaných molekul cholesterolu z krve a tkání, v jejich esterifikaci a v následném přenosu do jater. HDL tak působí proti rozvoji aterosklerózy, chrání LDL částice před oxidativními procesy a též stimuluje produkci oxidu dusnatého v cévních stěnách.
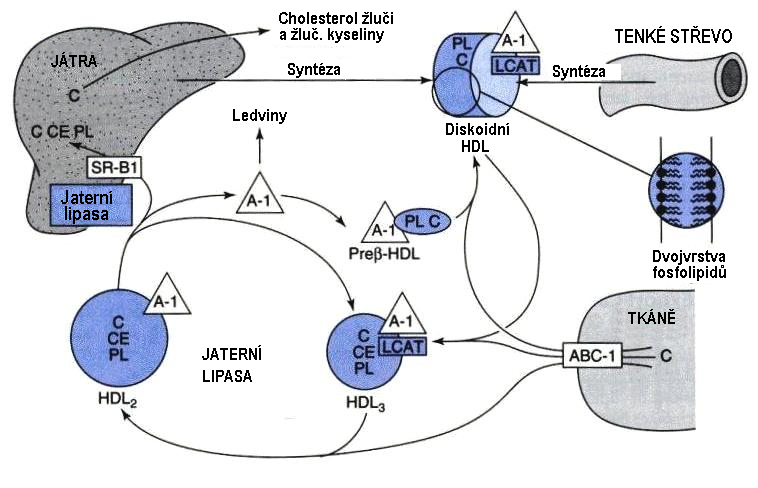
Obr. 10. Vznik a metabolismus lipoproteinů frakce HDL (upraveno z Murray et al. 2003)
Částice HDL jsou nositeli antiaterogenních účinků, které se realizují několika mechanismy:
- zpětným transportem cholesterolu z arteriální stěny do jater
- antioxidačními účinky
- protizánětlivými účinky; zlepšením endotelové funkce
- antiagregačním působením na trombocyty.
Za antioxidační účinky HDL jsou zodpovědné enzymy paraoxonasa 1 (PON1) a acetylhydrolasa faktoru aktivujícího destičky (platelet-activating factor acetylhydrolase – PAF-AH), které jsou součástí HDL. Paraoxonasa hydrolyzuje oxidované polynenasycené mastné kyseliny v pozici sn-2 fosfolipidů v oxidovaných LDL a tím chrání LDL před lipoperoxidací. PAF-AH se rovněž podílí na degradaci oxidovaných fosfolipidů, ale na rozdíl od PON1 působí na mastné kyseliny s nižším počtem uhlíků (C ≤ 9).
Protizánětlivé účinky se projevují snížením exprese adhezních molekul (např. VCAM-1, ICAM-1) na endotelu a inhibicí adheze monocytů k endoteliálnímu povrchu.
Lipoprotein (a)
Lipoproteiny Lp(a) představují třídu lipoproteinových částic, jejichž bílkovinnou složku tvoří apolipoprotein B-100 a specifický glykoprotein apolipoprotein (a), připojený k apoB‑100 disulfidovou vazbou (Obr. 11).
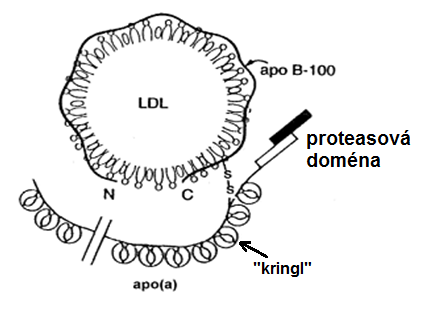
Obr. 11. Struktura lipoproteinu (a) (Jayasinghe et al. 2014)
Apoprotein (a) je syntetizován v játrech a na LDL částice se zřejmě váže v krevním řečišti. Fyziologická funkce apoproteinu (a) ani celého lipoproteinu (a) není známa. Ani chybění lipoproteinu (a) v plazmě nezpůsobuje žádnou chorobu ani syndrom z jeho deficitu. Uvádí se, že lipoprotein (a) podporuje transport lipidů do endotelu. Hyperlipoproteinémie Lp(a) je samostatným rizikovým faktorem aterosklerózy kardiovaskulárního aparátu, neboť vykazuje významnou nezávislou korelaci s onemocněním koronárních cév.
Základní jednotkou apoproteinu (a) je proteinová doména obsahující asi 80 aminokyselinových zbytků, vzájemně spojených třemi vnitřními disulfidickými můstky, čímž dochází k vytvoření zvláštního tvaru, který dává doméně její přívlastek “kringle”, tj. “preclík“ (prý podle podoby s typem dánského pečiva; český preclík vypadá podobně, to ale norský badatel Kåre Ingmar Berg při popisu struktury lipoproteinu (a) zřejmě nevěděl). Molekulová váha apolipoproteinu (a) se může u jeho četných polymorfních variant velice lišit a je dána právě počtem “kringlů”. Apoprotein (a) vykazuje značnou strukturní homologii s plasminogenem. Plasminogen je serinová proteasa, která je aktivována na plasmin, katalyzující trombolýzu. V molekule apoproteinu (a) je serin však zaměněn za arginin, takže Lp(a) je nefunkčním homologem plasminogenu. Apoprotein tak plasminogenu konkuruje – brání jeho aktivaci na plasmin a znemožňuje vazbu plasminu na fibrin. Za zvlášť významné se pokládá, že jeden z “kringlů“ v apo(a) se vyskytuje v mnoha tandemových kopiích. Čím více kringlů, tím výraznější je zásah apoproteinu (a) do plasminového fibrinolytického působení.