Faktory vyvolávající nádorovou transformaci
Vznik mutací, tedy nevratných kvalitativních nebo kvantitativních změn v genetické informaci, může být indukován fyzikálními, chemickými nebo biologickými faktory. Je třeba si uvědomit, že mutagenicita se automaticky nerovná karcinogenitě. Ačkoliv je vysoká pravděpodobnost, že mutageny jsou rovněž karcinogenní (~ 84 %), může docházet k falešným predikcím, protože i řada nekarcinogenních látek jsou mutageny a naopak celá řada nemutagenních látek může být karcinogenní. Řada látek a faktorů funguje jako kokarcinogeny tím, že zvyšují hladiny buněčných enzymů aktivujících karcinogeny.
Fyzikální onkogenní faktory
Mezi fyzikální faktory způsobující vznik nádorů patří ionizující záření (RTG, γ-záření), neionizující záření (UV A, UV B a UV C záření), chronické mechanické dráždění a vysoká teplota.
Ionizující záření (RTG, γ-záření) působí na DNA přímo a nepřímo. Při přímém působení dochází ke zlomům řetězců DNA, v jejichž důsledku vznikají chromosomové aberace, zkřížené vazby („cross-links“) či lokální denaturace (rozpojení) DNA. Při nepřímém působení reaguje ionizující záření nejprve s molekulami vody za vzniku ROS, které následně poškozují biomolekuly včetně DNA. Účinek závisí na dávce záření a na době působení. Nejohroženější je lymfatická tkáň.
Neionizující UV záření s delší vlnovou délkou (UV A) vyvolává tvorbu ROS, zatímco UV záření s krátkou vlnovou délkou (a tedy vyšší energií – UV B a UV C) reaguje přímo s molekulami DNA. UV B a UV C záření působí jako katalyzátor v reakci dvou sousedních thyminů v dvoušroubovici DNA za vzniku tzv. dimerů thyminu. Úsek DNA, který obsahuje takovýto dimer, představuje překážku pro DNA-polymerasu a nemůže být úspěšně replikován.
Chronicky působící vysoká teplota a chronické mechanické dráždění se uplatňují patrně jako promotory nádorových onemocnění díky chronické iritaci. Dlouhodobé pití horkého čaje (> 70°C) osminásobně zvyšuje riziko vzniku nádorů jícnu proti usrkávání čaje (< 65°C). Vdechnutí azbestu (vlákna < 60 nm) způsobuje mechanické poškození chromosomů s následnou fibrózou plic, bronchogenním karcinomem nebo maligním mezotheliomem.
Chemické onkogenní faktory
Podle epidemiologických studií je 75-80 % malignit způsobeno faktory životního prostředí, u 30-40 % obyvatelstva industrializovaných zemí se během života objeví zhoubný nádor. Faktory zevního prostředí, které se na rozvoji nádorů podílejí, jsou výživa (30 %), kouření tabáku (30 %), nadměrná konzumace alkoholu (10 %), profesionální expozice (5-10 %) a faktory lékové, průmyslové exhalace a další.
Chemické karcinogeny můžeme klasifikovat na karcinogeny genotoxické, epigenetické (negenotoxické) a kompletní (současně působí genotoxicky i epigeneticky). Genotoxické karcinogeny jsou látky s afinitou k DNA, které vytvářejí s nukleotidy kovalentní vazbu (vnesení mutace). Tyto látky zahajují maligní transformaci v somatických buňkách a jejich účinek je obvykle ireverzibilní. Teoreticky stačí k vyvolání účinku jediná expozice. Typický je elektrofilní charakter těchto látek, který mohou mít některé látky již primárně (přímé karcinogeny), častěji jsou však přítomny v neaktivní formě (prokarcinogen) a je nutná jejich metabolická aktivace na aktivní karcinogen s elektrofilními vlastnostmi (nepřímé karcinogeny). Epigenetické karcinogeny nereagují přímo s genetickým materiálem, ale usnadňují vznik nádorového onemocnění jinými mechanismy (např. imunosupresivními, hormonálními, cytotoxickými). Uplatňují se obvykle až v pozdějších stádiích karcinogeneze (většinou ve stádiu promoce).
Mezi chemické karcinogeny patří látky ze skupin polycyklických aromatických uhlovodíků, aromatických aminů, azo-barviv, nitrosaminů a nitrosamidů, hydrazo- a azoxysloučenin, karbamátů, halogenovaných sloučenin, přírodních produktů, anorganických karcinogenů a ostatních sloučenin (např. alkylační činidla, aldehydy, fenolické sloučeniny). Vzhledem k množství prokázaných chemických karcinogenů uvedeme dále jen příklady působení některých vybraných látek.
Polycyklické aromatické uhlovodíky (PAH) jsou látky obsahující systém kondenzovaných aromatických kruhů, nejvyšší riziko představují 17-PAH odvozené od fenantrenu (např. benzo[a]pyren, koronen, antracen, fluoren). Jsou to látky těžce rozpustné ve vodě, mají lipofilní charakter a koncentrují se v sedimentech. Vznikají zejména nedokonalým spalováním tuhých paliv, v dopravě, při kouření cigaret a při tepelné úpravě potravy (grilování, smažení, opékání na ohni). PAHy nevykazují akutní toxicitu, ale mohou poškodit celou řadu organismů, zejm. díky navození karcinogeneze a mutace DNA, poruchám reprodukce, podráždění až popálení pokožky. Jsou velmi odolné vůči přirozeným rozkladným procesům, kumulují se zejm. v rostlinách s velkou listovou plochou. Tyto sloučeniny jsou prokarcinogeny, které jsou v organismu biotransformovány na aktivní karcinogen působením různých monooxygenas, zejm. některými isoformami cytochromu P450 (CYP). Příkladem může být biotransformace benzo[a]pyrenu (BAP, prokarcinogen), který je nejprve pomocí CYP1A1 a CYP1B1 aktivován na BAP-7,8-epoxid, epoxidhydrolasa pak katalyzuje trans-adici vody za vzniku BAP-trans-7,8-diolu. Tento diol podstoupí další biotransformaci CYP1A1 nebo CYP1B1 za vzniku BAP-7,8-diol-9,10-epoxidu (BPDE), který je ultimátním karcinogenem schopným vytvářet kovalentní vazbu s guaninovými bazemi DNA (Obr. 7).
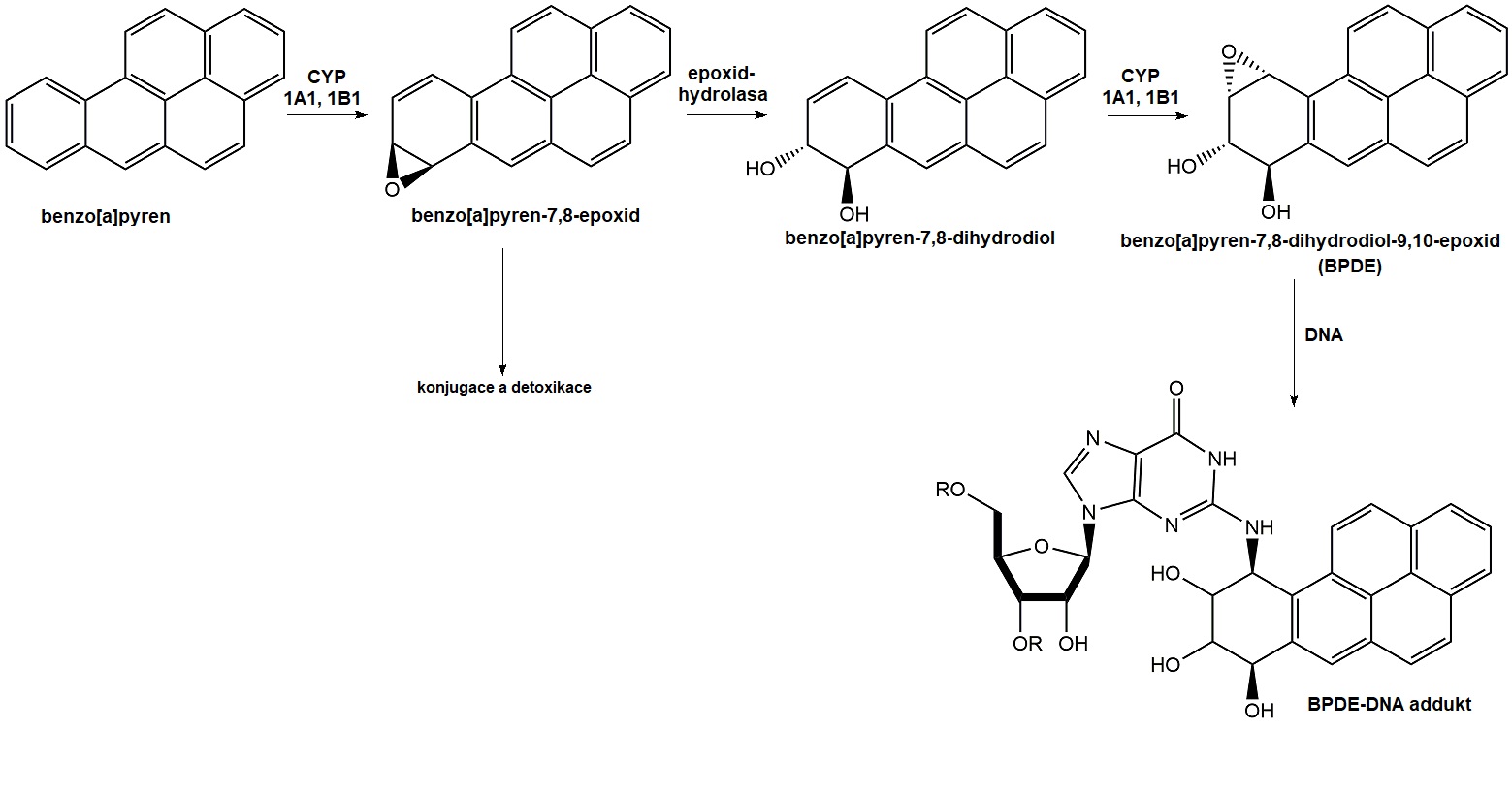
Obr. 7. Metabolické aktivace benzo[a]pyrenu a jeho vazba na DNA
Aromatický amin α-naftylamin byl po mnoho desetiletí používán v barvířství a barvířském průmyslu. Jedná se však o silný karcinogen s latencí 15-20 let. Samotný α‑naftylamin karcinogenní není, ale jeho biotransformací jaterními monooxygenasami vznikají toxické hydroxylované metabolity N‑hydroxy-2-naftylamin a 1-hydroxy-2-naftylamin, které jsou konjugovány s kyselinou glukuronovou na nekarcinogenní glykosidy. V močovém měchýři jsou hydrolýzou uvolněny aktivní karcinogeny, které vyvolávají vznik karcinomu močového měchýře (Obr. 8).
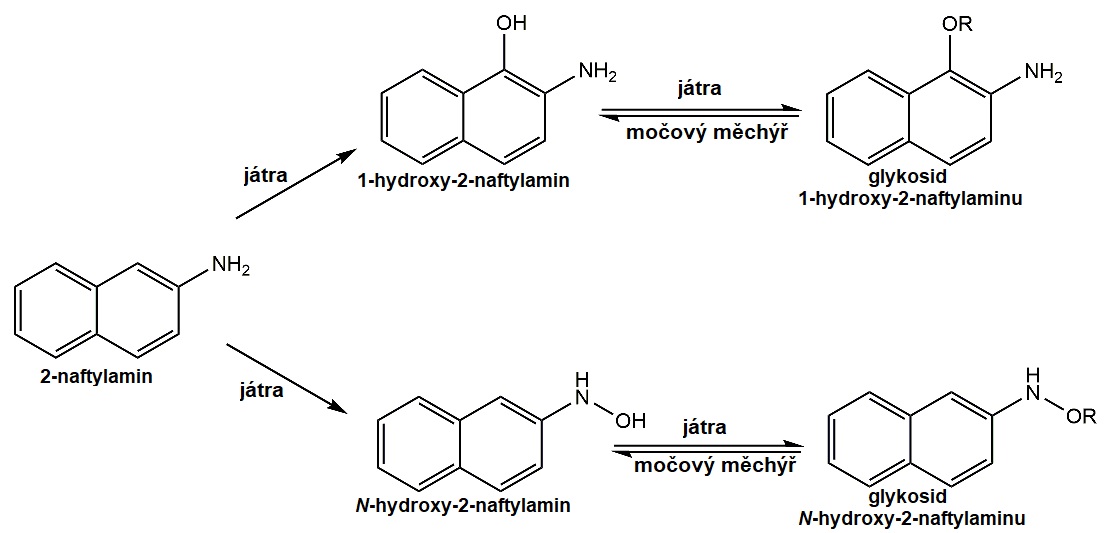
Obr. 8. Metabolická aktivace α-naftylaminu.
Karcinogeny potravy se jednak vyskytují přirozeně (např. rostlinné alkaloidy, toxiny) a lze se jim tedy snadno vyhnout, vznikají při tepelné úpravě potravy (PAHy, Maillardova reakce, pyrolýza proteinů a aminokyselin za vzniku heterocyklických aromatických aminů), jsou do potravy záměrně přidávány (např. potravní aditiva, barviva) nebo je jimi potravy kontaminována, a pak je velmi obtížné se jim vyhnout. Maillardova reakce je neenzymová reakce mezi redukujícími cukry (nebo jejich rozkladnými produkty) a volnými aminoskupinami proteinů v potravě. Tento proces je typický pro potraviny upravované pečením. Produktem jsou senzoricky aktivní látky, které poskytují potravě charakteristickou chuť, barvu (zlatohnědé zbarvení) a chuť, označované jako koncové produkty pokročilé glykace (AGE). Jedná se o komplexní směs stovek sloučenin, které nejsou ještě zcela charakterizované. Při vysokých teplotách však vznikají i karcinogenní látky, jako jsou akrylamid a 5-hydroxymethylfurfural. Neenzymová glykace však probíhá i v organismu (viz. diabetes mellitus).
U potravních aditiv (náhradní sladidla, umělá barviva, antioxidanty, dusičnany a dusitany) je často jen podezření na karcinogenní účinky. Dusičnany jsou nejprve nitroreduktasami v ústní dutině redukovány na dusitany. K redukci dusičnanů dochází i v žaludku při zvýšeném pH (nedostatek HCl) působením reduktas z ústní dutiny a reduktasami střevních bakterií při jejich průniku do žaludku. Dusitany pak reagují se sekundárními aminy potravy (sýry, ryby atd.) za vzniku nitrosaminů, které jsou mutagenní, karcinogenní a teratogenní. Nitrosaminy způsobují methylaci DNA a tím vyvolávají nádory jícnu a žaludku.
Rovněž některé přírodní složky potravy jsou zdrojem karcinogenů a/nebo mají mutagenní účinek. Obvykle však tyto efekty vykazují až ve vysokých koncentracích. Můžeme zde zmínit některé rostlinné polyfenolické látky (flavonoidy, třísloviny, lignany atd.). Tyto látky mají schopnost buď přímo interagovat s DNA, nebo zvyšují karcinogenní aktivaci jiných látek (např. flavonoid kvercetin a nádory močového měchýře). Safrol, látka ze skupiny fenylpropenů přítomná v rostlinách rodu Sassafras, je metabolizován na 1-hydroxyderivát, který je ve vyšších koncentracích hepatokarcinogenní. Pyrrolizidinové alkaloidy z podbělu, brutnáku či kostivalu mají hepatotoxické a hepatokarcinogenní účinky. V některých houbách (např. ucháč obecný, pečárky) jsou přítomné mykotoxiny s hydrazinovou skupinou (gyromytrin, agaritin), které po hydrolýze na aktivní karcinogen (substituovaný hydrazin) vyvolávají karcinomy plic nebo žaludku. Dalšími mykotoxiny s karcinogenními účinky jsou produkty plísní rodu Aspergillus, Penicillium a Fusarium – aflatoxiny, ochratoxiny, trichoteceny, zearalenony a fumonisiny. Člověk je jim vystaven při konzumaci kontaminované potravy (např. ořechů napadených plísní). Aflatoxin B1 je aktivován CYP 1A2 a CYP3A4 na toxický epoxid, který se váže na guaninové zbytky a vytváří tak addukty s DNA (Obr. 9). Karcinogenita tohoto metabolitu je způsobena vnesením bodové mutace do kodonu 249 genu kódujícího p53, kde dochází ke změně pořadí bází z AGT na AGG s následnou inaktivitou vznikajícího proteinu p53 a vznikem hepatocelulárního karcinomu.
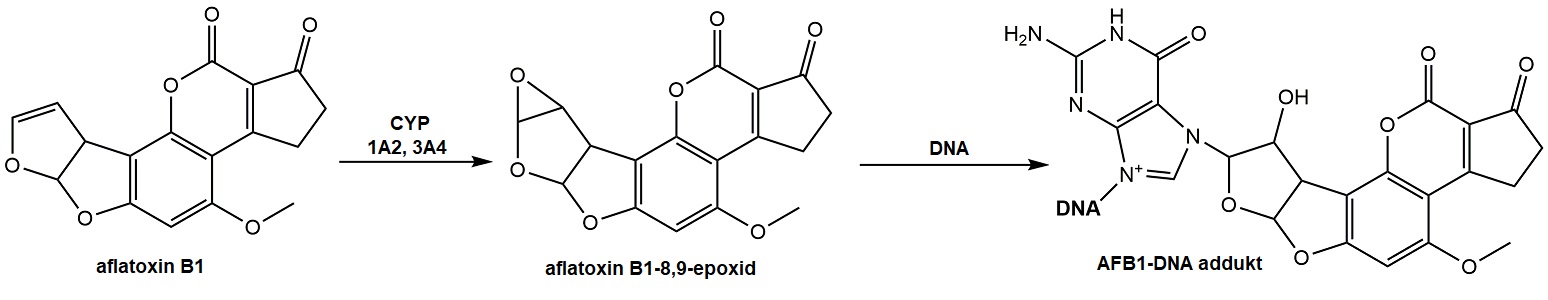
Obr. 9. Metabolická aktivace aflatoxinu B1 a jeho vazba na DNA
Biologické onkogenní faktory
U zhruba 20 % lidských nádorů se předpokládá, že jsou infekčního původu. Za 5-6 % z nich jsou zodpovědní parazité a bakterie. Někteří parazité, jako jsou např. motolice z rodu Schistosoma, se spolupodílejí na vzniku karcinomu močového měchýře jeho chronickou iritací. Drážděním se na vzniku karcinomu v otevřené kaverně podílí rovněž bakterie Mycobacterium tuberculosis. Některé nádory žaludku vznikají v důsledku antigenní stimulace při infekce Helicobacterm pylori, kdy se v žaludku vyvine lymfatická tkáň, která zde normálně není přítomna. Pokud antigenní stimulace přetrvává, dojde k lymfoidní hyperplazii s následným rozvojem maligního lymfomu žaludku z pozárodečných B-buněk, které již prodělaly antigenní stimulaci a selekci (MALT[1] lymfom).
Nejčastějším biologickým onkogenním faktorem jsou viry, u nichž se předpokládá, že se podílejí na vzniku zhruba 15 % lidských maligních nádorů. Virové onkogeny působí jako iniciátory nebo promotory karcinogeneze. Podmínkou onkogenity viru je integrace části nebo celého virového genomu do genomu hostitelské buňky. Studium onkogenních virů přispělo k porozumění principům maligní transformace. Byly popsány dvě třídy onkogenních virů – DNA viry a RNA viry (retroviry), které se liší mechanismem replikace.
DNA viry neobsahují ve svém genomu onkogeny. Onkogenní účinek těchto virů je zprostředkován jednak inaktivací tumor-supresorových genů hostitelské buňky a také inaktivací regulátorů buněčného cyklu virovými proteiny (posouvají hostitelskou buňku do S-fáze buněčného cyklu). Inaktivace p53 patří ke klíčovým událostem při maligní transformaci buňky. Příkladem onkogenního DNA viru je virus SV 40 (Simian Virus), vyvolávající lytickou infekci u opic. Genom viru SV 40 má dvě oblasti – časnou a pozdní. Časná oblast se exprimuje hned po infekci a je nezbytná pro syntézu virové DNA. Časná oblast kóduje dva proteiny – malý a velký T-antigen, které jsou klíčové pro replikaci viru a expresi hostitelského genomu (buněčná DNA-polymerasa). Pozdní oblast se exprimuje až po zahájení replikace virové DNA a zahrnuje geny kódující pro strukturní složky virové částice. V permisivních buňkách se podle lytického programu tvoří nové virové částice, buňka naplněná viriony následně lyzuje a zaniká, nemůže být proto transformována (plné uplatnění časných i pozdních virových genů). V non-permisivních buňkách je reprodukce viru zastavena, virová DNA v hostitelské buňce perzistuje volně nebo je integrována do hostitelského genomu (malá pravděpodobnost). Hostitelská buňka pak exprimuje specifické virové geny, které vyvolávají transformaci buňky na buňku nádorovou.
Mezi nejvýznamnější DNA-viry vyvolávající nádorovou transformaci u člověka patří viry z rodin Papovaviridae, Herpesviridae a Hepadnaviridae. Papillomaviry, které patří do rodiny Papovaviridae, jsou původci cervikálního karcinomu, dlaždicobuněčného karcinomu ústní sliznice a papilomatosy hrtanu. Lidské herpetické viry (rodina Herpesviridae) zahrnují lidský herpesvirus 8, spojovaný s Kaposiho sarkomem, a virus Epsteina a Barrové (EBV), který se podílí na vzniku Burkittova lymfomu, Hodgkinova a T-buněčného lymfomu a nasofaryngeálního karcinomu. Virus hepatitidy B (HBV, Hepadnaviridae) je spojován se vznikem hepatocelulárního karcinomu, jeho působení však bude pravděpodobně nepřímé a multifaktoriální.
Retroviry (RNA viry s reverzní transkriptasou) se mohou podílet na vzniku nádorů více mechanismy. Při replikaci retroviru v hostitelské buňce je nejprve virová RNA přepsána reverzní transkriptasou (RNA-dependentní DNA-polymerasa) do dvouvláknové DNA, která je následně inkorporována do genomu hostitelské buňky jako tzv. provirus. Primerem pro reverzní transkriptasu je tRNA z kapsidy retroviru. Následné transkripce jsou již zajištěny enzymovým aparátem hostitelské buňky. Retrovirus je nejvyšší parazit, protože jeho genom je stabilně spojen s genomem hostitelské buňky, vznikají nové viry, ale hostitelská buňka neumírá. Na základě biologické aktivity mohou být retroviry rozděleny do dvou skupin – pomalu a rychle transformující. Rychle transformující retrovirus obsahuje proti původnímu „typickému“ (pomalu transformujícímu) retroviru onkogenní sekvenci navíc (Obr. 10).
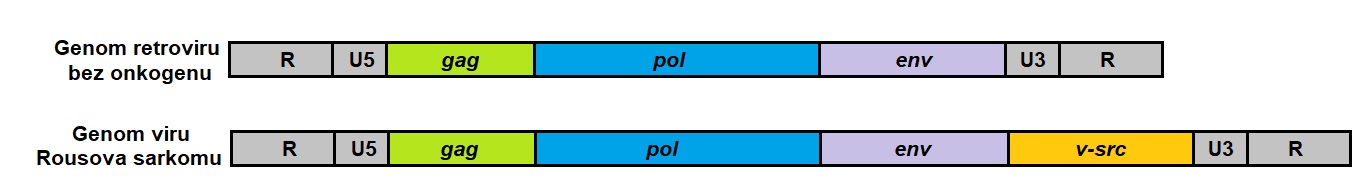
Obr. 10. Genom pomalu a rychle transformujícího retroviru
Pomalu transformující retroviry obsahují ve své RNA geny gag, pol a env, které zajišťují replikaci virové částice (gag kóduje antigenní proteiny kapsidy, pol virovou reverzní transkriptasu a env strukturní proteiny kapsidy a virového obalu). Proteinové produkty těchto genů nemají transformační účinek na hostitelskou buňku. Tyto viry neobsahují ve své genetické informaci onkogen, ale naruší inzercí své sekvence do genomu buňky sekvenci některého z protoonkogenů a tím jej aktivují (inzerce vysoce aktivního genového promotoru způsobí zvýšení jeho transkripce). Maligní transformace buňky po infekci těmito viry je pomalejší než u rychle transformujících virů a projevuje se až po dlouhé době latence (např. leukemie). U člověka byl transformující účinek prokázán pouze u lidského T‑lymfotropního viru (HTVL-1 a HTVL-2), který vyvolává T-buněčnou leukemii/lymfom dospělých.
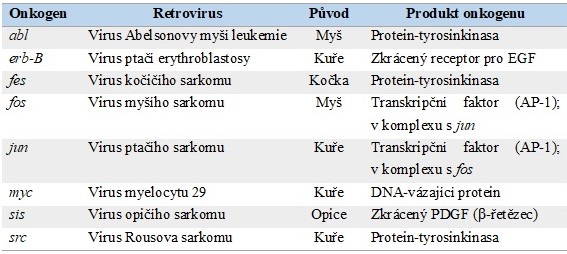
Rychle transformující retroviry obsahují ve své RNA oblast zajišťující replikaci viru (geny gag, pol a env) a navíc ještě některý z onkogenů (např. src, abl, fos, jun, myc), který vyvolá nádorovou transformaci hostitelské buňky. Tento onkogen je homologní s hostitelským protoonkogenem a dostal se do kapsidy při chybném vyštěpení vložené virové sekvence z buněčného genomu při resyntéze virových částic v průběhu infekce buňky. Inzercí této genetické informace do dalších infikovaných buněk dojde k amplifikaci sekvence určitého onkogenu v buňce a k rychlejšímu nástupu maligní transformace. Rychle transformující retroviry měly značný význam pro objevení buněčných (proto)onkogenů, ale vyskytují se pouze u zvířat. Příkladem je virus Rousova sarkomu způsobující maligní transformaci buněk u kura domácího, který obsahuje gen v‑src homologní s buněčným genem c-src. Příklady dalších retrovirových onkogenů jsou uvedeny v Tab. 2.
Tab. 2. Příklady onkogenů retrovirů (upraveno z Rodwell a kol. 2015)
[1] MALT – slizniční lymfatická tkáň (z ang. Mucosa Associated Lymphoid Tissue)