Prolomení antioxidační ochrany organismu, oxidační stres
V živém organismu je vznik RONS a jejich odstraňování antioxidačním systémem v přibližné rovnováze. Převaha jedné či druhé složky vede k poruchám, které mohou organismus vážně ohrozit. Častější je případ, kdy převládají RONS, ať je to způsobeno jejich zvýšenou tvorbou, sníženou činností AOX systému, nebo kombinací obou příčin. Při porušení této rovnováhy ve prospěch RONS vzniká stav, který se označuje termínem oxidační stres. I výrazná převaha AOX může mít nepříznivé následky, protože blokuje fyziologické a pro organismus nezbytné účinky RONS.
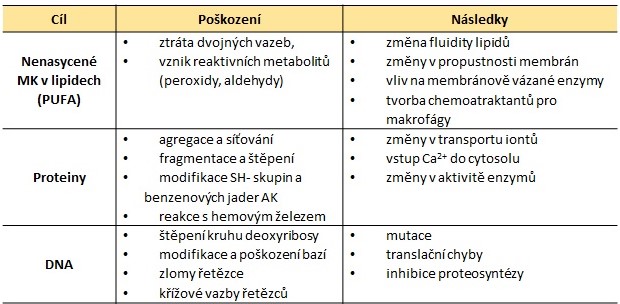
Prakticky kterákoliv biomolekula může být napadena a poškozena RONS. Nejzávažnější je poškození fosfolipidů biomembrán, vedoucí k poruše esenciálních membránových dějů či dokonce k zániku buňky, dále poškození nukleových kyselin (mutageneze, karcinogeneze, zánik buňky) a proteinů (inaktivace enzymů a dalších významných proteinů) (Tab. 4).
Tab. 4. Hlavní buněčné cílové struktury pro RONS (upraveno ze Štípek 2000)
Peroxidace lipidů (lipoperoxidace)
V biologických systémech jsou nejčastěji napadeny polynenasycené mastné kyseliny (PUFA). Peroxidace lipidů (Obr. 4) je typickým příkladem řetězové reakce, kde lze rozlišit iniciaci, propagaci a terminaci. Proces začíná (iniciace) vytržením vodíkového atomu z methylenové skupiny PUFA působením HO•, RO• nebo ROO• a mastná kyselina/lipid se stává uhlíkovým radikálem, v němž dvojná vazba přesmykuje tak, že vytváří konjugovaný dien. Ten se snadno konjuguje s kyslíkem za vzniku peroxylového radikálu (ROO•), který je dostatečně reaktivní, aby byl schopen napadnout další molekulu PUFA a vytvořit z ní radikál (propagace). Z původních ROO• vznikají hydroperoxidy (ROOH), které se cyklizují. Může dojít k rozštěpení řetězce cyklického peroxidu a uvolnění alkanů (etan, pentan), které vydechujeme, a reaktivních aldehydů, zejm. malondialdehydu (MDA) a 4-hydroxynonenalu (4-HNE). Řetězová reakce končí vznikem stabilní sloučeniny (terminace), např. při setkání radikálu PUFA s jiným radikálem nebo s lipofilním antioxidantem (zejm. α-tokoferol, koenzym Q10).
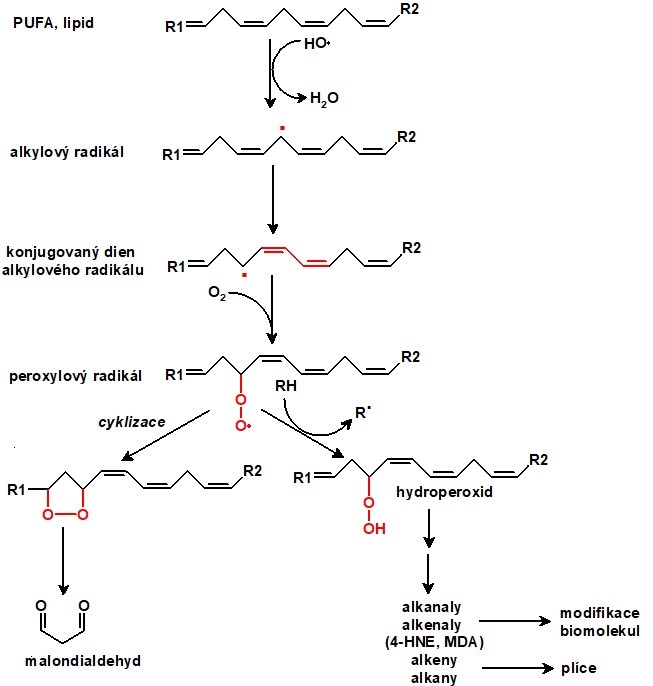
Obr. 4. Schéma peroxidace polynenasycené mastné kyseliny (upraveno ze Štípek 2000 a Racek 2003)
V organismu probíhají dva typy lipoperoxidace – enzymová a neenzymová. Enzymová lipoperoxidace probíhá v aktivních centrech hydro- a endoperoxidas (cyklooxygenasa, lipoxygenasa) a jejím výsledkem je vznik stereospecifických a biologicky aktivních látek – eikosanoidů. Volné radikály, které při jejich syntéze vznikají, však nejsou z enzymu uvolňovány, takže neškodí. Naproti tomu neenzymová lipoperoxidace je vyvolána nespecifickým a mnohdy patologickým podnětem a probíhá výše zmíněným mechanismem. Vznikající reaktivní aldehydy MDA a 4-HNE se pevně vážou na proteiny a mění jejich funkci a životnost. Výsledkem tohoto procesu jsou změny ve fluiditě membrán, vedoucí ke zvýšení jejich propustnosti pro ionty, což vyvolává lýzu buněk a snížení membránového potenciálu. Peroxidy vzniklé v membráně mohou být redukovány na příslušné alkoholy působením GPx, nebo mohou být z membrány odstraněny působením fosfolipas.
Poškození proteinů
Proteiny mohou být při oxidačním stresu poškozeny buď přímo – působením RONS, nebo nepřímo – vlivem produktů lipoperoxidace. Některé RONS jsou schopné bezprostředně oxidovat aminokyselinové zbytky, např. methionin je oxidován na methioninsulfoxid a následně methioninsulfon působením HO•, H2O2, 1O2, ONOOˉ a HOCl; tyrosinové, fenylalaninové a tryptofanové zbytky jsou nitrovány peroxynitritem či hydroxylovány pomocí HO•; oxidace prolinových a argininových zbytků je provázena vznikem glutamát-5-semialdehydu atd. Oxidace jednotlivých aminokyselinových zbytků není provázena řetězovou reakcí, jako tomu bylo v případě lipoperoxidace. V případě oxidace kostry polypeptidu však řetězová reakce probíhá a může dojít i k rozštěpení polypeptidu. Nepřímé poškození proteinů je způsobeno produkty lipoperoxidace (např. RO• a ROO•). Reaktivní aldehydy MDA a 4-HNE se kovalentně vážou na volné aminoskupiny lysinu za vzniku Schiffovy báze. Tato reakce vede k tvorbě příčných vazeb mezi sousedními řetězci proteinů, což způsobuje jejich agregaci a síťování. Při reakci 4-HNE s histidinovými, lysinovými či cysteinovými zbytky vznikají karbonylové sloučeniny, které jsou ukazatelem míry oxidačního poškození proteinů.
Cílem oxidačního poškození jsou nejen enzymy, ale i receptory a transportní proteiny. Následky těchto reakcí jsou závažné – enzymy mění svou aktivitu, iontové pumpy neudrží iontovou homeostázu buňky (v cytoplasmě se hromadí ionty Ca2+, které aktivují proteasy, fosfolipasy a endonukleasy), modifikace aminokyselinových zbytků způsobují vznik nových antigenních determinant. Vzniklé poškození proteinů může být částečně opraveno – disulfidové můstky jsou redukovány pomocí glutathionu a thioredoxinu na thiolové skupiny; methioninsulfoxid v proteinech je redukován zpět na methionin enzymem methioninsulfoxidreduktasou. Za in vivo podmínek bývají poškozené proteiny rovněž odstraněny proteolytickou degradací, avšak silně oxidované a agregované proteiny jsou vůči ní často rezistentní a v buňce se hromadí.
Poškození DNA
DNA je sama o sobě velmi stálá molekula, ale při jejím vystavení HO• vzniká celá řada oxidačních produktů, protože HO• napadá purinové a pyrimidinové baze i cukerné zbytky. Nejnáchylnější k oxidačnímu poškození je guanin. Častými produkty oxidačního poškození purinových bazí jsou 8-hydroxyguanin, 8-hydroxyadenin, 2,6-diamino-4-hydroxy-5-formamidopyrimidin (FAPyG) a 4,6-diamino-5-formamidopyrimidin (FAPyA), zatímco z pyrimidinových bazí vznikají cis- a trans-thyminglykoly, 5-hydroxy-5-methylhydantoin, 5‑(hydroxymethyl)uracil, cytosinglykol a 5,6-dihydroxycytosin. Při absorpci UV záření (200-300 nm) jsou pyrimidinové baze v DNA excitovány a při reakci se sousední pyrimidinovou bazí vytvářejí dimery (nejčastěji thyminové), které jsou chemicky stálé. RNS deaminují adenin na hypoxanthin, guanin na xanthin a cytosin na uracil, což může být příčinou chybného párování bazí a vzniku tranziční mutace. Cukerné zbytky deoxyribózy jsou vlivem HO• oxidovány a fragmentovány, přičemž vzniká celá řada produktů včetně karbonylových sloučenin a MDA. Následkem těchto reakcí může docházet k přerušení řetězce DNA (jedno- a dvouřetězcové zlomy) nebo se mohou vytvářet příčné vazby mezi DNA a proteiny.
Následkem oxidačního poškození DNA dochází k chybnému párování bazí při replikaci a zavedení chyby do genetické informace (tranzice v případě deaminace bazí pomocí RNS nebo v přítomnosti thyminglykolu, transverze v přítomnosti 8-hydroxyguaninu). Některé vznikající produkty blokují replikaci DNA (např. FAPyA či thyminglykol). Poškození DNA se projeví apoptózou, mutagenezí, karcinogenezí a stárnutím. Vzhledem k významu DNA pro přežití organismů existuje několik typů DNA reparačních enzymů – přímé odstranění změn (DNA-fotolyasa) a systémy pro excizní úpravy (excinukleasový systém, DNA-glykosylasa). Dimery thyminu i dalších bazí, které vznikají po ozáření DNA UV světlem, jsou rozštěpeny DNA-fotolyasou, která je závislá na energii viditelného světla. Chybně spárované, oxidované a deaminované baze jsou z DNA odstraněny buď pomocí excinukleasového komplexu, který při větších lézích vyštěpí poškozenou bazi jako součást oligonukleotidu, nebo je poškozená baze odstraněna přímo pomocí DNA-glykosylasy. Prázdná místa v DNA jsou pak opravena a doplněna pomocí endonukleasy, DNA-polymerasy a DNA-ligasy.