Fyziologické funkce RONS
Ve většině publikací je zdůrazňován nepříznivý účinek RONS. Nesmíme však zapomenout, že RONS se účastní celé řady pochodů, které jsou pro přežití buňky/organismu nezbytné. Škodí pouze tehdy, vymknou-li se přísné kontrole, kterou každý aerobní organismus získal v průběhu vývoje biologického systému.
Metabolismus živin a zisk energie
Poslední částí dýchacího řetězce mitochondrií je cytochrom-c-oxidasa (komplex IV), která přijímá elektrony pocházející z živin a redukuje jimi molekulární kyslík. Molekula kyslíku je postupně čtyřmi elektrony redukována na dvě molekuly vody a při tomto procesu se uvolní energie pro syntézu ATP. Během redukce vznikají v aktivním centru reaktivní metabolity (viz. Obr. 1), které zůstávají navázány na enzym. Vzhledem k nepředvídatelným vlastnostem ROS je však zhruba 3-5 % denní spotřeby kyslíku přeměněno na O2ˉ•, H2O2, HO• a 1O2 místo kompletní redukce na vodu.
Monooxygenasy, enzymy endoplasmatického retikula, redukují kyslík třemi elektrony na HO•, který využívají k hydroxylačním reakcím, např. při syntéze cholesterolu a žlučových kyselin, při detoxikaci některých xenobiotik (cytochromy P450), při degradaci tryptofanu kynureninovou cestou (indolamin-2,3-dioxygenasa).
Další enzymy, které využívají kyslík, jsou různé oxidasy. Jednou z nich je xanthindehydrogenasa, která oxiduje hypoxanthin a xanthin na kyselinu močovou. Prolyl- a lysylhydroxylasy jsou enzymy syntézy kolagenu, které využívají molekulární kyslík k posttranslační hydroxylaci prolinových a lysinových zbytků za účasti kofaktorů askorbátu a 2‑oxoglutarátu. Tyrosinhydroxylasa, která má jako kofaktor tetrahydrobiopterin, zahajuje syntézu katecholaminů zavedením hydroxylové skupiny do pozice 3 na aromatickém jádře tyrosinu.
Při syntéze hormonů štítné žlázy dochází k oxidaci jodidu peroxidem vodíku na elementární jód, který je následně využit k jodaci tyrosinových zbytků v proteinu thyreoglobulinu. Tyto reakce katalyzuje specifická peroxidasa – thyreoperoxidasa, zdrojem peroxidu vodíku jsou duální oxidasy DUOX1 a DUOX2, které patří mezi NADPH-oxidasy.
Deoxyribonukleotidy vznikají in vivo redukcí ribonukleosiddifosfátů v přítomnosti enzymu ribonukleotidreduktasy (též ribonukleosiddifosfátreduktasa). Tento enzym má v aktivním místě přítomný RS•, který se účastní katalýzy reakce. Pro redukci 2-OH skupiny ribosy se využívá pár thiolových skupin (cysteinových zbytků) v aktivním místě enzymu, který se během reakce oxiduje. Regenerace enzymu probíhá pomocí thioredoxinového nebo glutaredoxinového systému za účasti NADPH jako donoru redukčních ekvivalentů.
Fagocytóza
PMN a makrofágy používají RONS k odstraňování mrtvých buněk a k zabíjení bakterií. Jejich membrána je vybavena enzymovým komplexem – NADPH-oxidasou, který po pohlcení cizí částice aktivuje a redukuje kyslík na O2ˉ•, čímž se 10-20 násobně zvýší spotřeba O2 aktivovanou buňkou („respirační vzplanutí“). Superoxid je superoxiddismutasou redukován na peroxid vodíku. Kyselé prostředí fagosomu podporuje uvolnění iontů železa z intracelulárních zásob, takže je k dispozici pro Fentonovu reakci a vznik HO•. U aktivovaných PMN je přítomná myeloperoxidasa, která katalyzuje vznik HOCl z peroxidu vodíku a chloridových iontů. V makrofázích i PMN je účinkem cytokinů, mikrobů a jejich produktů stimulována exprese indukovatelné NO-syntasy, která (v závislosti na dostupnosti kofaktorů, substrátu a NADPH) ve svém okolí zvyšuje koncentraci NO o několik řádů. Ten pak nestačí být odstraněn difúzí do krve a reaguje ve fagosomu se superoxidem za vzniku baktericidního prostředku ONOOˉ (Obr. 2).
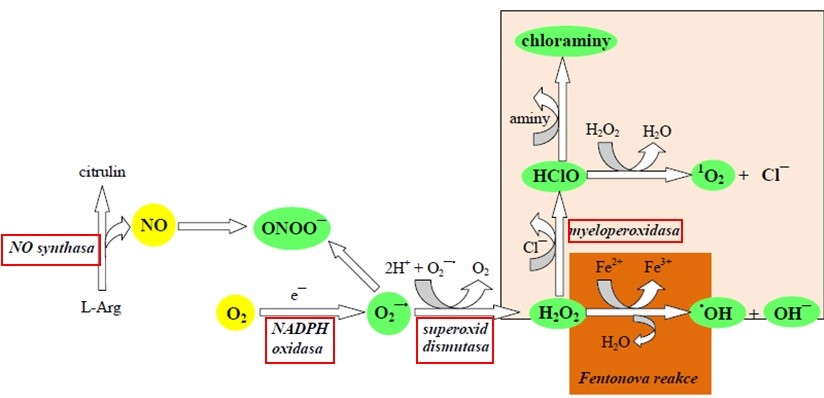
Obr. 2. Tvorba RONS ve fagocytujících buňkách (upraveno z Krejsek a kol. 2016)
Buněčná signalizace
Indukce tvorby RONS (pouze nízké koncentrace!) významně ovlivňuje aktivitu drah signální transdukce. RONS mohou hrát do určité míry úlohu druhých poslů v signálních drahách aktivovaných cytokiny a růstovými faktory. Mezi nejvýznamnější cíle RONS patří modulace aktivity různých transkripčních faktorů (např. Nrf2, AP-1, NF-κB, HIF-1), proteinkinas (např. MAPK, PI(3)K) a fosfatas. Výsledkem je např. stimulace buněčné proliferace, stárnutí (senescence) buněk, modulace apoptózy atd. Významnou signální molekulou je NO, který působí jako neurotransmiter a vazodilatační faktor. Tento radikál aktivuje guanylátcyklasu, která v buňce syntetizuje cGMP.