Volné radikály, reaktivní formy kyslíku a dusíku
Volné radikály jsou látky (atomy, molekuly, ionty) schopné samostatné existence, které mají ve svém elektronovém obalu nepárový elektron (ev. více nepárových elektronů). Vznikají z normálních částic ztrátou elektronu (oxidací), přijetím elektronu (redukcí) či homolytickým štěpením kovalentní vazby, při kterém každý fragment získá jeden nepárový elektron. K homolytickému štěpení je třeba značné množství energie (např. ultrafialové nebo ionizační záření, teplota ~ 450-600 °C), proto vznikají volné radikály v biologických systémech většinou energeticky snadnějším způsobem (ztrátou či přijetím elektronu). Volné radikály mohou být neutrální částice, kladně nebo záporně nabité ionty v závislosti na tom, zda počet protonů v atomovém jádře odpovídá počtu elektronů v orbitalech (neutrální částice) nebo neodpovídá (ionty). Vzorce radikálů se označují tečkou, která indikuje nepárový elektron, a vzorec je doplněn podle počtu a typu náboje znaménkem + či -, pokud je popisovaná částice zároveň iontem.
Homolytické štěpení kovalentní vazby A-B → A• + B•
Redukce (molekula přijímá elektron) A + eˉ → Aˉ•
Oxidace (molekula ztrácí elektron) B → B+• + eˉ
Volné radikály jsou obvykle málo stabilní a vysoce reaktivní, protože se snaží doplnit si chybějící elektron a dosáhnout tak stabilní elektronové konfigurace. Vznik radikálu může být iniciací celého řetězce dalších reakcí. Chybějící elektron získává volný radikál vytržením z „normální“ molekuly, ze které se tak stává radikál a radikálová reakce se tím propaguje do okolí. Radikálová reakce je ukončena (terminace) buď reakcí dvou radikálů, nebo reakcí s látkou, jejíž radikál je stabilní a může existovat delší dobu (např. α-tokoferol či jiné nízkomolekulární antioxidanty).
Pojem reaktivní formy kyslíku (ROS) či reaktivní formy dusíku (RNS) je souhrnným označením pro volné radikály a sloučeniny spojené s metabolismem kyslíku nebo dusíku, které nemají nepárový elektron (např. peroxid vodíku či peroxynitrit). V Tab. 1 jsou uvedeny příklady nejvýznamnějších RONS spolu s jejich řádovým biologickým poločasem.
Tab. 1. Reaktivní formy kyslíku a dusíku s řádovým poločasem některých z nich (upraveno podle Štípek 2000)
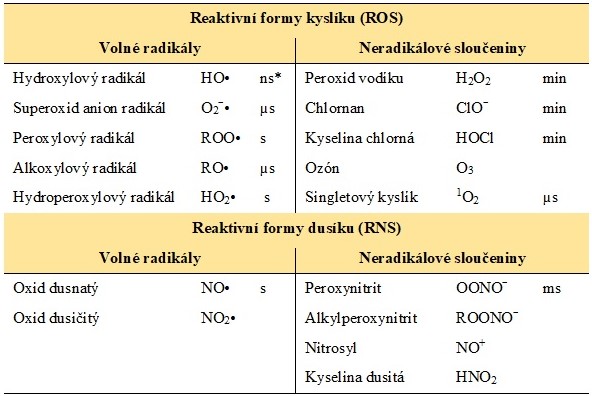
*řádový biologický poločas dané RONS v živém organismu
Reaktivní formy kyslíku
V organismu se nejčastěji setkáváme s volnými radikály odvozenými od kyslíku. Běžnou dvouatomovou molekulu kyslíku (též tripletový dioxygen) můžeme rovněž zařadit mezi radikály, protože ve dvou molekulových orbitalech obsahuje po jednom nepárovém elektronu (je to biradikál), které mají stejné spinové kvantové číslo. Ve srovnání s jinými radikály však reaguje s biomolekulami poměrně pomalu, protože příjem elektronu vyžaduje, aby jeden ze stávajících nepárových elektronů změnil svůj spin (rotaci), což je relativně pomalý proces, a překonal tak tzv. spinovou restrikci. Tato „drobnost“ umožnila živým organismům přežít v kyslíkové atmosféře planety Země, jinak by doslova shořeli. Molekulární kyslík se však může snadno změnit v reaktivnější formu tím, že mu dodáme energii nebo jinak porušíme spinovou restrikci (změnou počtu elektronů).
Při postupné redukci molekuly kyslíku čtyřmi elektrony vznikají postupně další ROS a nakonec dvě molekuly vody (Obr. 1). Přijetím prvního elektronu vzniká superoxid anion radikál (O2ˉ•), což je vůbec nejhojnější volný radikál v živých organismech. Superoxid se přijetím dalšího elektronu redukuje na peroxid vodíku (H2O2), který se v přítomnosti dalšího elektronu rozpadá na vodu (disociovaná forma hydroxidový ion HOˉ) a hydroxylový radikál (HO•), což je vůbec nejreaktivnější volný radikál v živých organismech s extrémně krátkým biologickým poločasem. Poslední elektron redukuje hydroxylový radikál na molekulu vody (opět ve formě HOˉ). Popsaná reakce probíhá např. v aktivním centru cytochrom-c-oxidasy (enzym dýchacího řetězce mitochondrií) a umožňuje finální transformaci energie chemických vazeb živin do ATP.
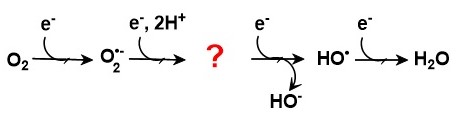
Obr. 1. Postupná čtyřelektronová redukce molekuly kyslíku
Superoxid má oxidační i redukční vlastnosti a podléhá dismutaci, při které jedna jeho molekula poskytuje elektron druhé, takže O2ˉ• se vlastně zároveň oxiduje i redukuje. Produkty reakce jsou kyslík a peroxid vodíku. Přestože reakce probíhá ve vodném prostředí velkou rychlostí, v organismu je ještě o čtyři řády urychlena enzymem superoxiddismutasou.
Z patobiochemického hlediska stojí za pozornost též singletový kyslík (1O2), který vzniká excitací jednoho elektronu tripletového kyslíku do stavu s vyšší energetickou hladinou, která je provázena změnou spinu tohoto elektronu. Tato změna uspořádání elektronů, díky které je odstraněna spinová restrikce, způsobí značný nárůst reaktivity. V organismu vzniká 1O2 při spontánní dismutaci superoxidu (jedna molekula superoxidu se oxiduje na kyslík a druhá se zároveň redukuje na peroxid vodíku) a po absorpci světla některými pigmenty (tzv. nepřímé fotosenzitizační reakce). Takovými pigmenty (fotosenzitizéry) jsou např. retinal, bilirubin či porfyriny, které jsou po absorpci světla o vhodné vlnové délce excitovány a mohou přenést excitační energii na sousední molekulu kyslíku, ze které se stává 1O2, a fotosenzitizér se vrací do základního energetického stavu. Singletový kyslík pak napadá buď molekulu fotosenzitizéru a/nebo jinou přítomnou molekulu, čímž se změní její struktura a vlastnosti (fotodynamický účinek). Některá onemocnění (např. porfyrie) jsou provázená excesivním vznikem 1O2. Fotodynamické reakce se však používají i k terapii, např. novorozenecké žloutenky, psoriázy či k fotodynamické terapii solidních tumorů.
Vznik kyseliny chlorné (HOCl) je katalyzován enzymem myeloperoxidasou, která je přítomná v azurofilních granulech neutrofilních granulocytů (PMN). Jedná se o silný oxidant, který PMN používají jako mikrobicidní prostředek. Při reakci HOCl s aminoskupinami proteinů vznikají chloraminy, které jsou stabilnější, ale mají slabší mikrobicidní účinky než HOCl.
Peroxylové (ROO•) a alkoxylové radikály (RO•), které vznikají např. během peroxidace lipidů, jsou dobrými oxidačními činidly. Alkoxylové radikály se však v organismu rychle přeměňují na jiné typy radikálů.
Vzájemné přeměny ROS jsou katalyzovány ionty přechodných kovů, zejména ionty železa a mědi. Jedná se o prvky z první řady d-prvků (železo, měď, mangan), které mají neúplně obsazenou předposlední valenční slupku elektronového obalu, proto velmi ochotně reagují s ROS. Většina jejich biologických účinků (benefičních i škodlivých) souvisí právě se schopností přijímat a darovat elektrony. V organismu jsou vázány v depotních formách (např. transportní a skladovací proteiny transferin, feritin, ceruloplasmin), kde jsou udržovány v redoxně inaktivním stavu. Příkladem vzájemné přeměny ROS jsou Haberova-Weissova reakce a Fentonova reakce. Haberova-Weissova reakce spontánně bez katalýzy neprobíhá, protože O2ˉ• ve vodném prostředí spontánně nereaguje ani s H2O2, ani s organickými hydroperoxidy. V přítomnosti železa však reakce H2O2 a O2ˉ• probíhá velmi snadno, a to ve dvou krocích. Nejprve O2ˉ• zredukuje železité ionty na železnaté a ty pak pohotově redukují H2O2 na HO•, který je vysoce toxický. Poslední reakce je známá jako Fentonova reakce a jedná se o dlouho známý děj a také nejčastější způsob vzniku toxického HO•.
Haberova-Weissova reakce (1934) H2O2 + O2ˉ• → O2 + HO• + HOˉ
Redukce Fe3+ O2ˉ• + Fe3+ → O2 + Fe2+
Fentonova reakce (1894) H2O2 + Fe2+ → HO• + HOˉ + Fe3+
Reaktivní formy dusíku
Molekulu oxidu dusnatého (NO) můžeme zařadit mezi volné radikály, protože obsahuje jeden nepárový elektron. Biologický význam NO je obrovský – je součástí procesů nespecifické imunity, působí jako neurotransmiter v CNS a VNS a ovlivňuje cévní tonus a tím reguluje krevní tlak (léčebné využití při Angině pectoris – NO se uvolňuje z nitroglycerinu či amylnitritu). Přestože se jedná o jednoduchou molekulu, syntetizuje se složitým mechanismem z aminokyseliny L-argininu za katalýzy enzymu NO-synthasy (NOS). Existují čtyři typy NOS, které se liší regulací exprese (konstitutivní x indukovatelná), závislostí na Ca2+ a primárním místem izolace (endoteliální x neuronální x makrofágová x mitochondriální). NO reaguje s většinou biomolekul velmi pomalu, protože jeho difúze do krve a inaktivace hemoglobinem probíhá mnohem rychleji, dostatečně rychle reaguje jen s jinými radikály a přechodnými kovy. Metabolity NO (např. peroxynitrit) jsou však velmi reaktivní. Při reakci s thiolovými skupinami cysteinu, glutathionu či albuminu vznikají stálejší nitrosothioly, které jsou zřejmě transportní formou NO.
Reakcí NO se superoxidem vzniká toxický peroxynitrit (ONOOˉ), který je silným cytotoxickým oxidačním agens. In vivo vyvolává hydroxylaci a nitraci tyrosinových zbytků a oxidaci methioninových zbytků v proteinech, což vede k inaktivaci enzymů (např. α1‑antiproteinasy, trombomodulinu, glutathion-S-transferas, cytochromů P450) a interferenci se signální transdukcí (zablokování fosforylace tyrosinových zbytků pomocí tyrosinkinas).
Sirné radikály
V in vivo podmínkách vystupují thiolové sloučeniny (zejm. redukovaný glutathion) většinou jako antioxidanty, protože chrání thiolové skupiny proteinů před oxidací a vychytávají kyslíkové radikály a další reaktivní formy (např. HOCl a ONOOH). Thioly (včetně glutathionu) však mohou být rovněž zdrojem radikálů (thiylové radikály, RS•). Thiylové radikály vznikají po reakci thiolů s radikálem lokalizovaným na uhlíku nebo kyslíku (HO•, ROO•, RO•), po reakci thiolů s ionty přechodných kovů nebo homolytickým štěpením disulfidových vazeb (i v proteinech). Thiylové radikály mají značnou reaktivitu, mohou oxidovat další látky za vzniku jiných radikálů (např. askorbylový radikál z askorbátu, O2ˉ•).