Opakování principů buněčné signalizace
Buňky mnohobuněčných organismů jsou vystaveny vnitřním i mimobuněčným signálům, na něž reagují. Tyto signály řídí růst, dělení a diferenciaci během vývoje, stejně jako chování buněk v dospělých tkáních. Jádrem všech těchto komunikačních systémů jsou chemické signály, které jsou vysílány jednotlivými buňkami a zachycovány zpravidla jinými buňkami, jimž jsou určeny.
Život buňky závisí na extracelulárních signálech, které buňka zachycuje prostřednictvím receptorů a reaguje na ně. Buňky různých typů se od sebe liší svou receptorovou výbavou, která umožňuje reagovat odpovídajícím způsobem na signální molekuly produkované jinými buňkami. Reakce buňky odpovídá tomu, jaká kombinace signálů na ni působí. Konkrétní jednotlivá buňka zpravidla vyžaduje k přežití více signálů současně. Další signály vyvolávají růst a dělení, jiné signalizují diferenciaci. Buňka, která nedostává potřebné signály k přežití, podstoupí formu buněčné sebevraždy známé jako apoptóza (Obr. 1). Skutečná situace je ještě složitější.
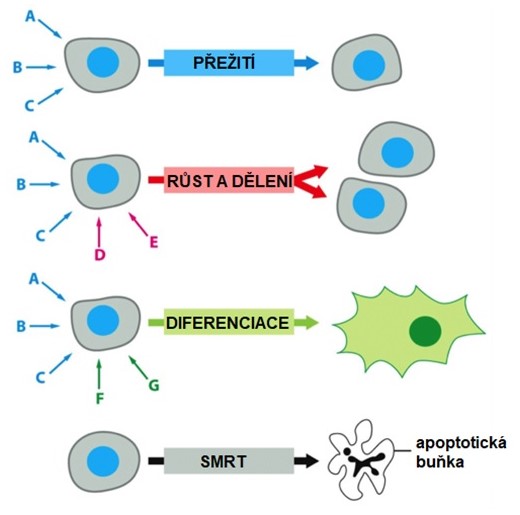
Obr. 1. Závislost živočišné buňky na příjmu extracelulárních signálů (upraveno z Alberts et al. 2007).
Mezibuněčná komunikace se uskutečňuje na různé vzdálenosti (Obr. 2). Např. existují komunikace sousedních buněk nebo buněk blízkých (parakrinní signály). Komunikace na velké vzdálenosti může probíhat prostřednictvím nervů (elektrochemická), nebo vysíláním signálů do tělních tekutin (zejména do krve – klasická endokrinní komunikace). Některé signály mohou být určeny vlastní buňce (autokrinní signály).
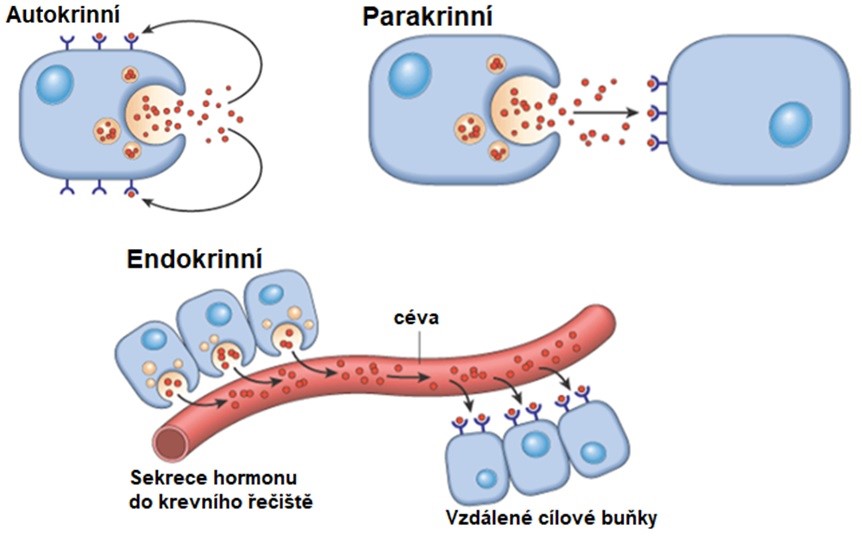
Obr. 2. Autokrinní, parakrinní a endokrinní komunikace (Kumar et al. 2014).
Většina buněk mnohobuněčných organismů signály vysílá i přijímá. Buňka, která vysílá signál, je buňka signální, buňka, která přijímá signál, je buňka cílová. Příjem signálů závisí na receptorových proteinech cílových buněk. Cílová buňka je opatřena příslušnými receptory pro konkrétní signály. Tyto receptory mohou být na povrchu buňky (membránové receptory) nebo uvnitř buňky (intracelulární receptory). Vazba signálu aktivuje receptor, který následně aktivuje jednu nebo více intracelulárních signálních drah nebo systémů. Tyto systémy závisí na intracelulárních signálních proteinech, které zpracovávají signál uvnitř přijímající buňky a distribuují jej do příslušných intracelulárních cílů. Na konci signálních drah jsou efektorové proteiny, které uskutečňují příslušnou změnu v chování buňky. V závislosti na signálu, typu a stavu přijímající buňky mohou tyto efektory fungovat jako regulátory transkripce, iontové kanály, složky metabolické dráhy nebo části cytoskeletu. Schéma endokrinní signalizace ilustruje Obr. 3.
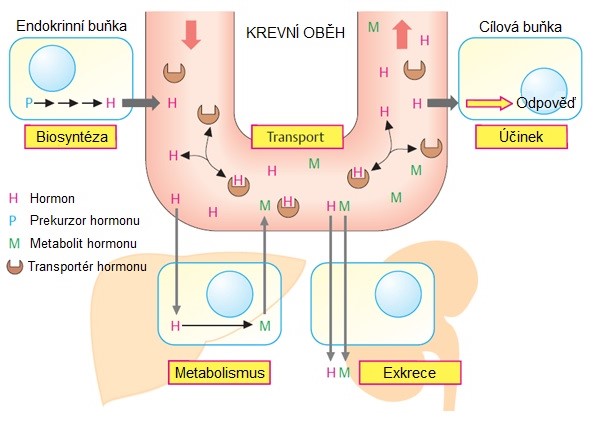
Obr. 3. Obecné schéma hormonální signalizace (upraveno z Koolman a Roehm 2005)
Signály vysílané buňkami do krve se tradičně nazývají hormony. Buňky, které produkují hormony, se nazývají buňky endokrinní. Tyto buňky tvoří endokrinní žlázy (viz Obr. 4), nebo jsou rozptýlené („dispersní žlázy“, např. v gastrointestinálním traktu).
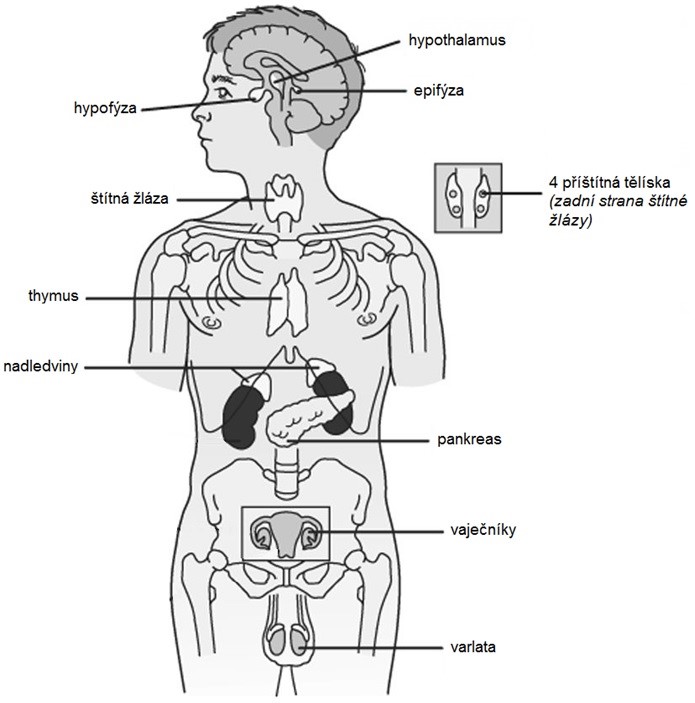
Obr. 4 Přehled žláz s vnitřní sekrecí (upraveno z Kalbasi 2015)
Chemická povaha hormonů a signálních molekul obecně je velmi pestrá. Známe hormony odvozené od aminokyselin (např. adrenalin, thyroxin), mastných kyselin (prostaglandiny), peptidové a proteohormony (např. insulin či glukagon) a hormony steroidní, které vznikají modifikací molekuly cholesterolu (např. kortisol, aldosteron či estradiol). Přehled hormonů podle struktury je v Obr. 5.
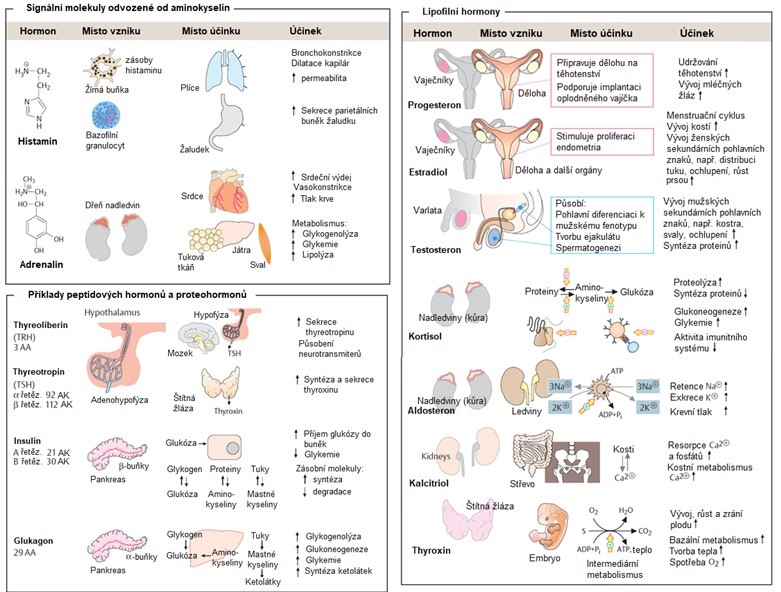
Obr. 5 Příklady hormonů, jejich struktury a funkce (upraveno z Koolman a Roehm 2005).
Chemická povaha hormonů a signálních molekul obecně je velmi pestrá. Chemická povaha hormonu zároveň určuje, jakým způsobem je hormon transportován krví a jak je přijímán cílovou buňkou. Z tohoto hlediska můžeme rozlišit dvě základní skupiny hormonů. Polární molekuly (proteiny, peptidy, hormony odvozené od aminokyselin…) jsou v krvi rozpuštěné a cílovou buňkou je signál přijat prostřednictvím příslušného povrchového – membránového receptoru. Hormony lipofilní (steroidní hormony, hormony štítné žlázy, …) jsou transportovány krví ve vazbě na více nebo méně specifický transportní protein. Pronikají buněčnou membránou cílové buňky a váží se na intracelulární receptor. Schéma dalšího postupu signálu cílovou buňkou je znázorněno v obr. 6 a 7.
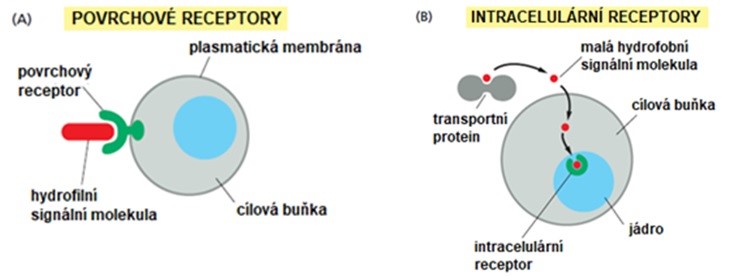
Obr. 6 Příjem hydrofilního a lipofilního signálu cílovou buňkou (upraveno z Alberts et al. 2007)
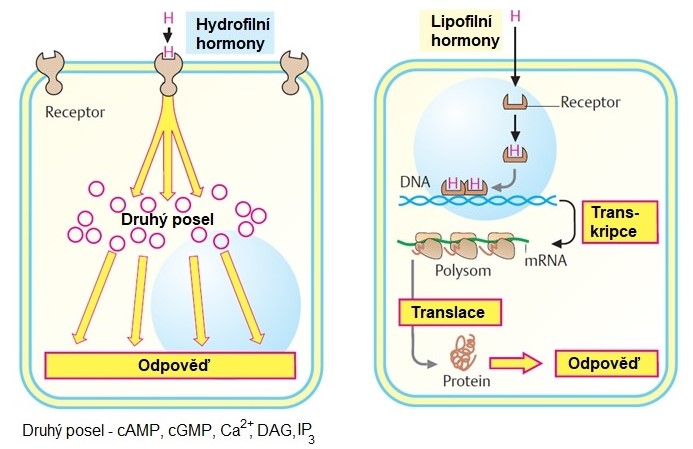
Obr. 7 Hydrofilní a lipofilní hormony. Principy převodu signálu v cílové buňce (upraveno z Koolman a Roehm 2005)
Jednou z charakteristik hormonálního působení je amplifikace signálu, modulace dalšími faktory a rozrůznění cílových efektů (Obr. 8).
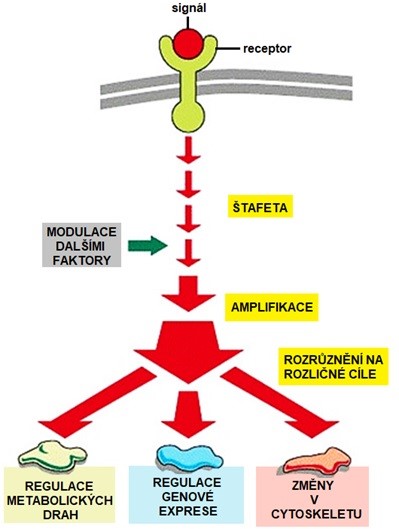
Obr. 8 Schéma intracelulární signalizace hydrofilního hormonu (upraveno z Alberts et al. 2007)
Základní typy membránových (povrchových) receptorů ukazuje Obr. 9.
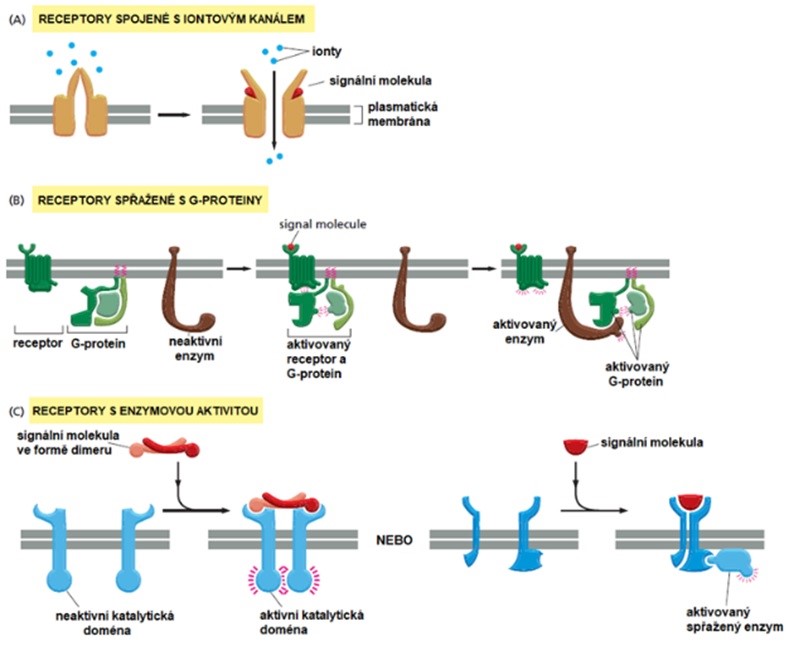
Obr. 9 Typy membránových receptorů pro hydrofilní signály (upraveno z Alberts et al. 2007).
Cílovým enzymem v případě signalizace prostřednictvím G-proteinu (obr. 10) může být např. adenylátcyklasa, jejímž produktem je cyklický AMP (cAMP) jako „druhý posel“, přenášející signál „prvního posla“, tj. hormonu, do nitra buňky. Jiným enzymem, který je aktivován prostřednictvím G-proteinu, je fosfolipasa C. Jejím substrátem je fosfatidylinositol-bisfosfát (PIP2) a produktem jsou dva „druzí poslové“ – inositol-trifosfát (IP3) a diacylglycerol (DAG). Cesta aktivace fosfolipasy C je spojena s účastí vápenatých iontů. Viz obr. 10 a podrobný výklad v Obecné biochemii.
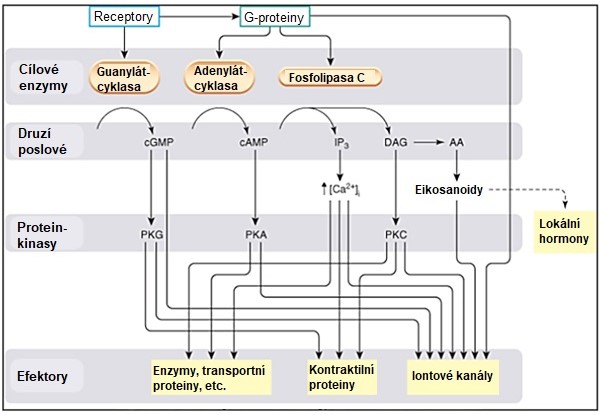
Obr. 10. Přenos signálu cestou G-proteinu (upraveno z Ritter et al. 2019)
U člověka existuje více než 800 různých G-proteinů, z nichž polovina zprostředkovává smyslové vjemy (chuť vůni, zrak)!
Existuje několik endokrinních poruch, způsobených mutacemi, jejichž výsledkem je ztráta nebo změna funkce příslušného G-proteinu nebo receptoru spřaženého s G-proteinem. Prvním popsaným příkladem takové poruchy byl pseudohypoparathyreoidismus, charakterizovaný renální rezistencí na parathormon.
Poznání signalizace cestou G-proteinu přispělo k porozumění způsobu, jak působí toxiny bakterií, vyvolávajících choleru a černý kašel (pertuse). Tyto toxiny mají enzymovou aktivitu, interferující s normálním přenosem signálu u hostitele. Cholera toxin, produkovaný bakterií Vibrio cholerae, se vyskytuje např. v kontaminované vodě, katalyzuje přenos ADP‑ribosy z koenzymu NAD+ na α-podjednotku Gs, čímž blokuje GTPasovou aktivitu této podjednotky a tím udržuje Gs trvale v aktivovaném stavu. Tím je stále permanentně aktivní adenylátcyklasa buněk střevního epitelu a tím vysoká koncentrace cAMP, který spouští stálou sekreci chloridových a bikarbonátových iontů a vody do střevního lumen. Výsledná ztráta vody a elektrolytů je hlavním příznakem cholery. V případě pertuse mikroorganismus Bordetella pertusis produkuje toxin pertuse, který katalyzuje ADP-ribosylaci Gi, čímž brání výměně GDP za GTP a blokuje inhibici adenylátcyklasy tímto G-proteinem.
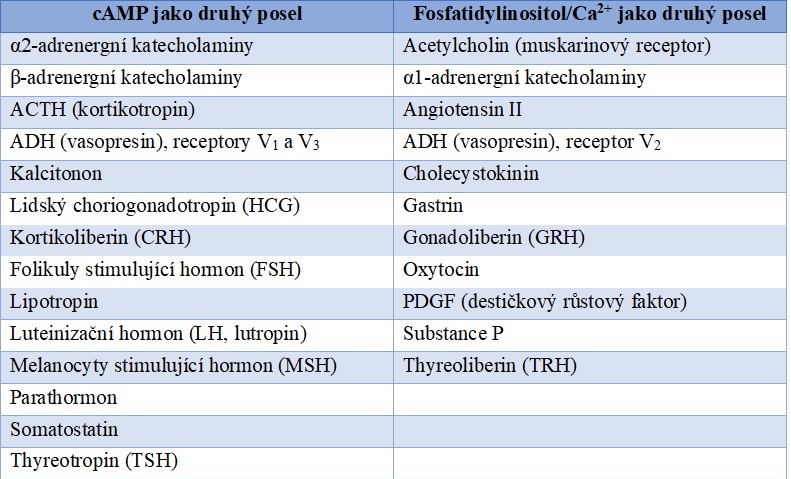
Tabulka 1 Příklady signálních molekul, jejichž intracelulární signalizace je zprostředkována G-proteiny
Některé signální dráhy jsou krátké a jejich zpětnovazebná regulace je jednoduchá, viz Obr. 11.
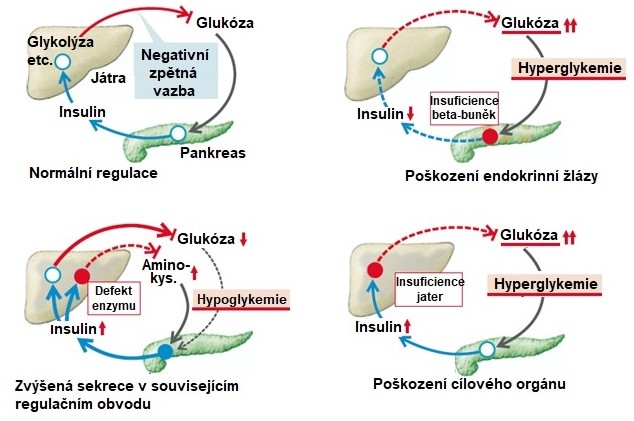
Obr. 11. Příklad jednoduchého endokrinního regulačního obvodu a jeho dysregulace (upraveno ze Silbernagl a Lang 2012).
Endokrinní signální dráhy jsou však v řadě případů kaskádové, a regulace hormonální aktivity probíhá na principu krátké nebo dlouhé zpětné vazby. Obr. 12 ukazuje tento typ regulace na příkladu osy „Signál - vyšší mozková centra - hypothalamus - hypofýza - periferní hormon“. Vícestupňový kaskádový hormonální systém ukazuje zároveň na uplatnění principu amplifikace signálu a negativních zpětných vazeb.
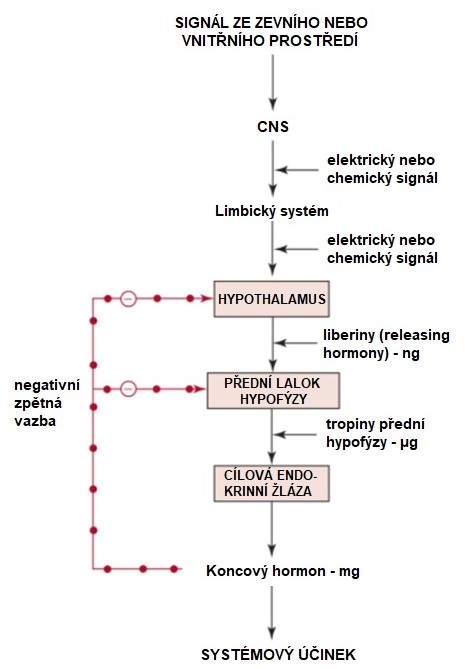
Obr. 12 Kaskádová hormonální signalizace a zpětnovazebné endokrinní regulace (upraveno z Devlin 2011)