Hemoglobin – struktura, funkce, poruchy globinové části molekuly
Hemoglobin
Hemoglobin (Hb) je červené krevní barvivo, které je součástí erytrocytů obratlovců a některých dalších živočichů.
Hemoglobin zabezpečuje přenos krevních plynů – především kyslíku z jeho zdroje do periferních tkání. Vedle toho se podílí na transportu oxidu uhličitého v opačném směru. Hemoglobin také tvoří důležitý pufrační systém krve, který odstraňuje přebytek H+ (zejména v periferních tkáních těla) vazbou na histidinové zbytky svých molekul. Hemoglobin je lokalizován v erytrocytech, v nichž tvoří asi 35 % jejich celkového obsahu.
Molekula hemoglobinu je sférického tvaru a skládá se ze 4 podjednotek. Každá podjednotka je tvořena bílkovinnou částí – globinem a prostetickou (nebílkovinnou) částí – hemem (Obr. 34).
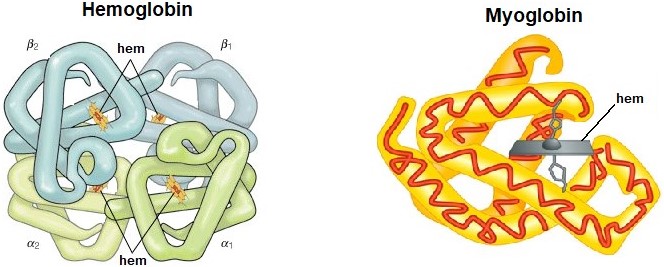
Obr. 34. Struktura hemoglobinu A (vlevo) a myoglobinu (vpravo). Každá ze čtyř podjednotek hemoglobinu obsahuje jeden hem (červené políčko), zatímco jednořetězcový myoglobin obsahuje pouze jeden hem (upraveno z The Editors of Encyclopaedia Britannica 2018 a Timberlake 2012).
Vždy dvojice polypeptidových řetězců je stejná. V lidském organismu se vyskytují řetězce alfa, beta, gama, delta atd. Na základě přítomnosti jednotlivých řetězců v molekule pak hovoříme o různých typech hemoglobinu, které se vzájemně liší afinitou k molekule kyslíku (schopnost vázání nebo uvolňování samotné molekuly O2).
Železo má v molekule hemu celkem 6 koordinačně kovalentních vazeb (koordinační číslo 6). Centrální atom železa ve formě Fe2+ je vázán čtyřmi z těchto vazeb k atomům dusíku v pyrrolových jádrech porfyrinu. Páté místo se váže na zbytek histidinu globinového řetězce. Šesté místo může být obsazeno molekulou kyslíku (Obr. 35).
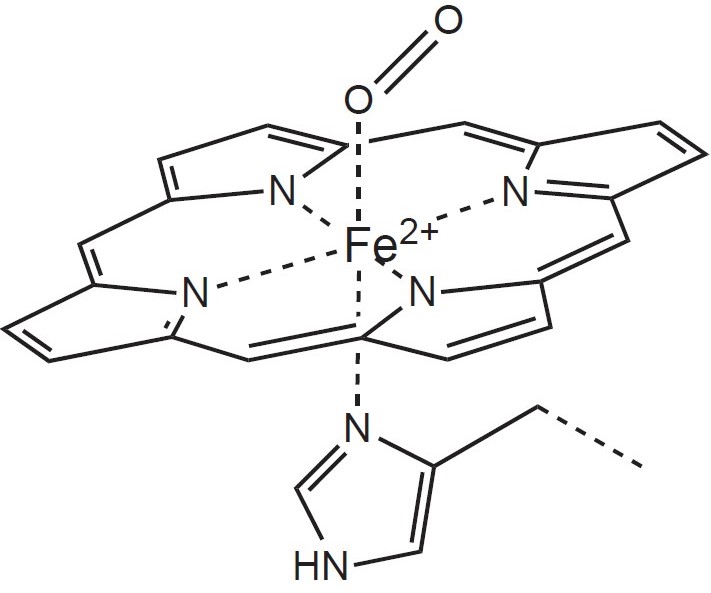
Obr. 35. Vazba kyslíku na hem v molekule hemoglobinu (Lavríková a Fontana 2018).
Kromě kyslíku se mohou na hem hemoglobinu vázat i jiné ligandy, jako je oxid uhelnatý (vysoká afinita, kompetice s kyslíkem). Vazbou halidu, kyanidu nebo hydroxidu se mění Fe2+ na Fe3+ a vzniká methemoglobin, který neváže ani nepřenáší kyslík (Obr. 36).
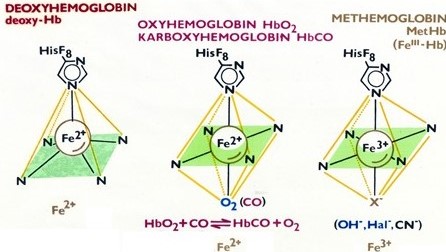
Obr. 36. Vazba kyslíku a alternativních ligandů na hemoglobin (Kolektiv autorů).
Vazba kyslíku mění vztahy mezi hemoglobinovými řetězci, a tedy i kvarterní strukturu molekuly (Obr. 37).
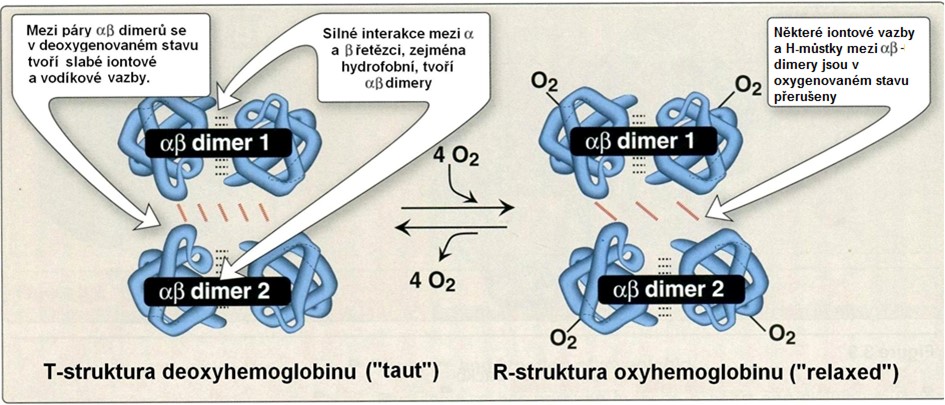
Obr. 37. Změna kvartérní struktury molekuly hemoglobinu při vazbě a uvolnění kyslíku (deoxyhemoglobin a oxyhemoglobin) (upraveno z Harvey a Ferrier 2011)
Při vazbě a uvolňování kyslíku se uplatňuje kooperace globinových řetězců, která je ukázkou kooperativního efektu molekul, složených z několika podjednotek. Grafické vynesení závislosti sycení hemoglobinu kyslíkem na jeho koncentraci (parciálním tlaku) poskytuje typickou sigmoidální křivku, rozdílnou od hyperboly, znázorňující vazbu kyslíku na jednořetězcovou molekulu myoglobinu s jedním hemem (Obr. 38).
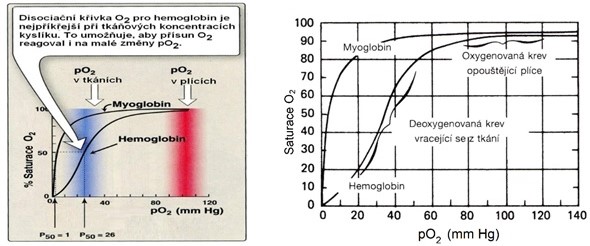
Obr. 38. Porovnání saturace myoglobinu a hemoglobinu kyslíkem (disociační křivka) (upraveno podle Harvey a Ferrier 2011 a Murray et al. 1998).
Z obrázku je zároveň zřejmé, že v plicích se hemoglobin plně sytí kyslíkem a v tkáních ho odevzdává myoglobinu.
Lokalizace genů pro globinové řetězce
Skupina (cluster) genů příbuzných α-genu je lokalizována na 16. chromosomu. Lokus pro α-globin je tetraplikován a gen pro ζ (zeta) globin je duplikován. Skupina (cluster) genů příbuzných β-genu je lokalizována na 11. chromosomu a je tvořena genem β a δ, genem γ G a γ A a genem ε (epsilon) (Obr. 39).
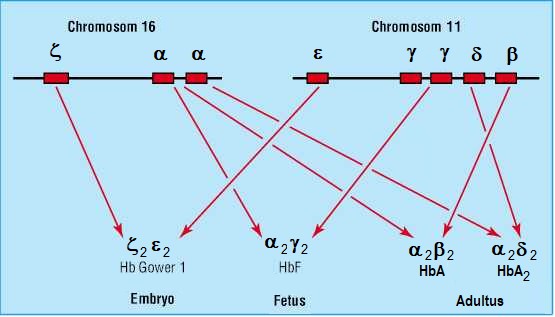
Obr. 39. Umístění genů pro jednotlivé řetězce na chromosomech (upraveno z Alauldeen 2015)
V průběhu života od embryonálního stádia přes období plodu a dále po celé další období života jedince po porodu se v souladu se změnou podmínek vystřídá ve své funkci několik typů hemoglobinu (Tab. 5).
Tabulka 5. Fyziologické typy hemoglobinu v průběhu ontogeneze (upraveno z Přispěvatelé Wikiskript 2019a)
Postupná změna složení hemoglobinu v průběhu ontogeneze odpovídá způsobu, jakým jedinec kyslík pro svůj život získává z prostředí. V embryonálním stádiu musí mít hemoglobin dostatečnou afinitu ke kyslíku, aby dostatečně přebíral a vázal kyslík, difundující z okolní tkáně matky (zásobené kyslíkem prostřednictvím jejího hemoglobinu), a dodával jej tkáním embrya. Fetální hemoglobin musí mít dostatečnou afinitu k převzetí kyslíku z placentálních klků, což je snadnější cesta. Ve srovnání s těmito typy má nejsnadnější úlohu hemoglobin dospělých HbA, který se sytí kyslíkem z bohaté nabídky v plicních alveolech při vysokém parciálním tlaku kyslíku. Všechny tyto typy hemoglobinu musí být zároveň ochotny předat navázaný kyslík tkáním, resp. myoglobinu. Tomu odpovídají relativně jiné saturační křivky různých typů hemoglobinu. Z obrázku je zřejmé, že samotné řetězce α nebo β nejsou schopny tuto funkci plnit. Jejich saturační křivka je obdobná, jako saturační křivka myoglobinu (Obr. 40).

Obr. 40. Sycení fyziologických typů hemoglobinu kyslíkem. (upraveno z Manning et al. 2017).
Embryonální hemoglobin je tvořen buňkami krevních ostrůvků žloutkového váčku v prvních týdnech vývoje. Rozeznáváme tyto typy embryonálního Hb: Gower I (2ζ2ε), Gower II (2α2ε), Portland 1 (2ζ2γ) a Portland 2 (2ζ2β).
V průběhu vývoje dochází ke změnám v expresi jednotlivých genů – tzv. přepínání (switching) globinů. Nejprve je zahájena syntéza zeta a epsilon globinů (HbGower 1). Následně dochází k expresi alfa a gama a globinů a vzniká tak fetální HbF; současně jsou suprimovány zeta a epsilon geny a ve fetálním období se tvoří převážně HbF. U novorozence obsahují erytrocyty si 70 % HbF; v dospělosti již jen 1 %. Regulace tvorby hemoglobinu v ontogenezi souvisí s lokalizací tvorby červených krvinek. Embryonální hemoglobin se tvoří ve žloutkovém váčku, fetální v játrech a dospělý v kostní dřeni (Obr. 41).
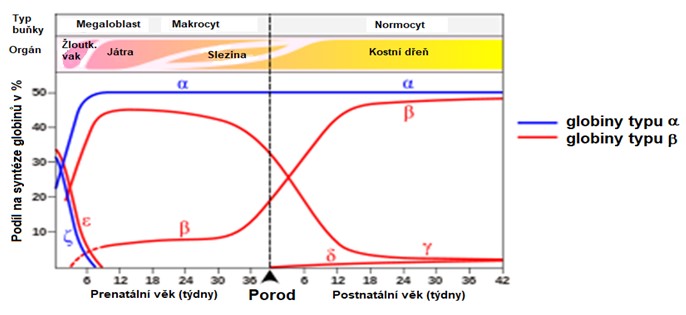
Obr. 41. Tvorba jednotlivých řetězců hemoglobinu v průběhu vývoje jedince (upraveno z Wood 1976).
Vazebná (disociační) křivka hemoglobinu je ovlivněna aktuálními podmínkami. Významným regulátorem je např. 2,3-bisfosfoglycerát (BPG), vznikající jako by-pass produkt při glykolýze.
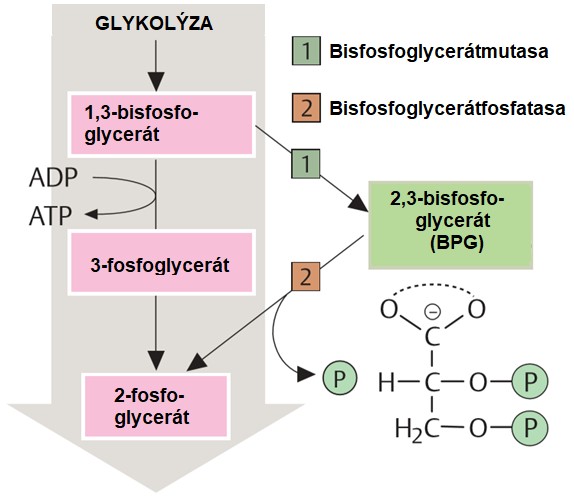
Obr. 42. Tvorba 2,3-bisfosfoglycerátu (upraveno z Koolman a Roehm 2005)
Bisfosfoglycerát je alosterickým regulátorem hemoglobinu. Stabilizuje deoxyhemoglobin, protože se váže svými čtyřmi záporně nabitými fosfáty na β-podjednotky hemoglobinu, na jejichž zrušení (tj. na oxygenaci Hb) je třeba dodat určité množství energie. Funkce BPG tedy spočívá v podpoře předání kyslíku v tkáních (Obr. 43).
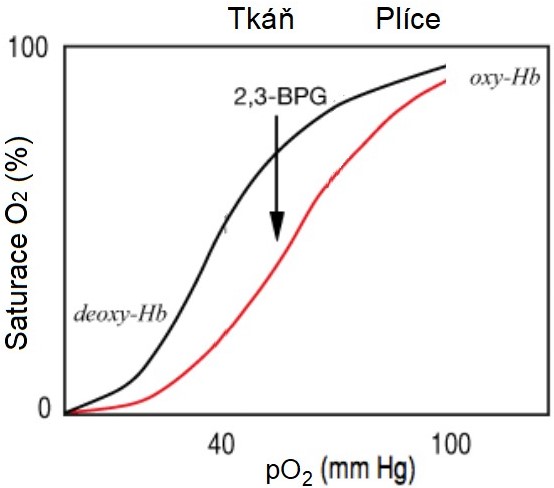
Obr. 43. Vliv 2,3-bisfosfoglycerátu na saturaci hemoglobinu kyslíkem (upraveno z David 2018)
V tkáních je pH nižší (při metabolismu vzniká CO2). Oxid uhličitý zde difunduje do erytrocytu, kde tvoří kyselinu uhličitou za katalýzy enzymem karboanhydrasou. Kyselina uhličitá disociuje, bikarbonát je pak transportován erytrocytem. V plicích, kde je naopak nízký parciální tlak CO2, probíhá opačný proces a uvolněný oxid uhličitý je vydýchán (Obr. 44).
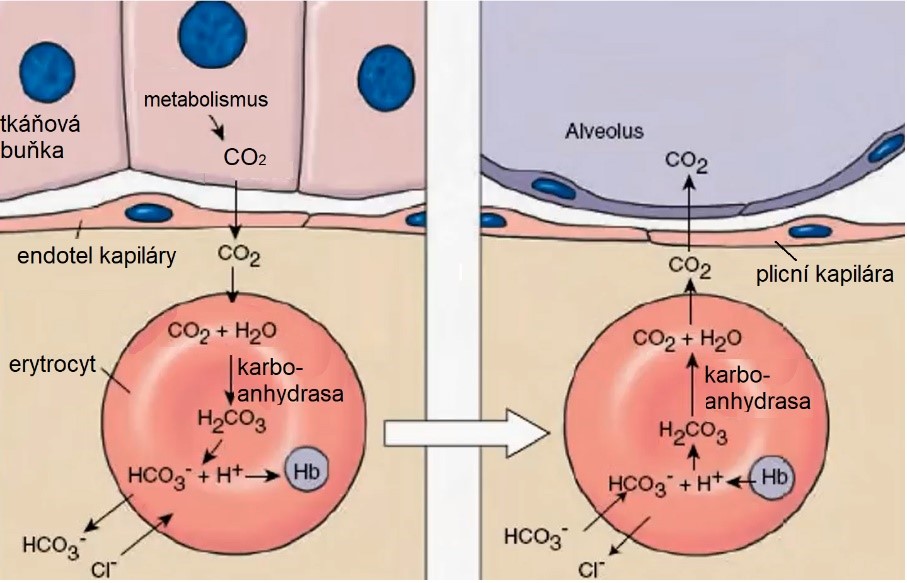
Obr. 44. Účast erytrocytu v transportu oxidu uhličitého z tkání do plic (upraveno z Kluthe 2014). Vyšší koncentrace CO2 znamená zároveň snížení pH.
Sycení hemoglobinu kyslíkem je ovlivněno také pH. Nižší pH při průtoku krve tkáňovými kapilárami proto působí ve prospěch uvolnění kyslíku z vazby na hemoglobin (Bohrův efekt) (Obr. 45).
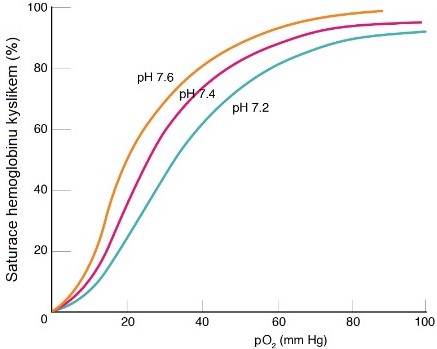
Obr. 45. Vliv pH na saturaci hemoglobinu kyslíkem (Bohrův efekt; upraveno Kolektiv autorů)
Při saturaci hemoglobinu kyslíkem se uplatňuje i vliv teploty. Čím vyšší teplota, tím nižší afinita hemoglobinu ke kyslíku a naopak, čím je teplota nižší, tím vyšší afinita hemoglobinu ke kyslíku (Obr. 46).

Obr. 46. Komplexní působení pH, koncentrace CO2, BPG a teploty na saturaci hemoglobinu kyslíkem (Přispěvatelé WikiSkript 2019b)